Transformed cells
Both DNA and RNA viruses cause tumors in animals. Both are able to transform mammalian cells in tissue culture. Transformed cells may be recognized by their following characteristics:
(1) multiplication forever (immortalization)
(2) grow to a higher saturation density
(3) have reduced requirement for serum
(4) grow in suspension in soft agar
(5) form different cell colony patterns
(6) grow on top of normal cell monolayers
(7) readily agglutinate by lectins
The transformed cells may cause tumors in animals but this is usually a several-stage process in which transformation is but the first step. In addition, tumor formation involves complex interactions with the immunological system of the host animal, hence transformation in culture does not necessarily mean malignancy in vivo. Transformation simply provides some clue to early events required for the development of malignant potential by infected cells. (BSc 2nd Year Microbiology Viruses Notes Study Material)
Oncogenesis. There is no single mechanism by which viruses induce tumors. The possible mechanisms, involving infection of tumor precursor cells, by which viruses may induce tumors are as follows.
(1) Persistence of all/part of the viral genome which
(a) expresses a viral gene (an oncogene) that initiates or maintains transformation
(b) alters the control or expression of cellular genes (insertional mutagenesis)
(c) requires immunosuppression.
(2) No evidence of prior infection (“hit-and-run).
For the study of oncogenesis two major theories have been predominant:
(1) That there is the deletion of genetic material essential for normal functioning, or
(2) That there is a very specific activation of genetic material which could be a positive or negative control element, either de novo or to an abnormal level.
This theory is also known as gene activation theory.
There is much information on oncogenes. There are cancer genes that were first identified in retroviruses and which have a counterpart in normal cells. The viral oncogenes are known as V-onc and the cellular progenitors from which they were derived as proto-oncogenes or Conc. The transformed cells are essentially normal cells that no longer have their normal control over proliferation or differentiation and, they require constant positive action for the transformed state to be maintained. (BSc Microbiology Viruses Notes Study Material)
Carcinogenesis. The full-blown cancer cell is not created at a stroke but is the result of a series of changes one or more may be due to the effect of a tumor virus. Carcinogenesis is multifactorial; various factors in addition to virus infection, such as host genotype, diet, environmental carcinogens (other than viruses), and other invading organisms (as HIV acts as a cofactor for the development of Kaposi’s Sarcoma by Cytomegalo virus) all contribute to the process. Transformation is one of the earlier stages en route to cancer.
In virus-induced carcinogenesis, the immune system is important as it provides a check on virus multiplication and the spread of cells bearing viral (tumor) antigens. It is not a major factor in non-viral carcinogenesis since animals with deficient immune responses do not develop more non-viral-induced tumors than normal animals.
Benign tumors, such as warts, are most certainly caused by viruses but still, we do not know with certainty if any human malignancies are of viral origin. Since we do not have full knowledge of the actual mechanisms involved in carcinogenesis, evidence implicating viruses in human cancer is still fragmentary. It should at the same time be surprising that viruses are not responsible for some human cancers since they induce different types of malignancy in several animals. Viruses appear to be involved in human cancers.
The best evidence links Epstein-Barr virus (EBV) with nasopharyngeal carcinoma, EBV with Burkitt’s lymphoma, hepatitis B virus with primary hepatic carcinoma, and cytomegalic virus with Kaposi’s sarcoma. Human papillomavirus is linked with cervical cancer. Definite proof of viral etiology of human cancer will only be obtained if Koch’s postulates could be fulfilled.
Oncogenic viruses
Various members of four different families listed in Table are known to be oncogenic in animals or are suspected of having oncogenic properties in man. All, except Retroviruses, are deoxyviruses. Even in Retroviruses, there is involved DNA intermediate in the replication of viral RNA. There are no ‘pure’ oncogenic riboviruses. What are Retroviruses?
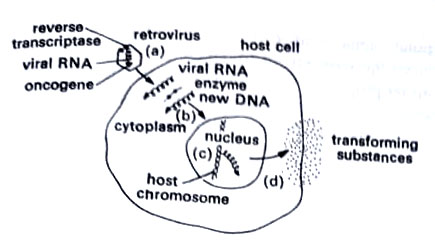
Retroviruses. They are so named because their reverse transcriptase “reverses” the flow of genetic information from RNA to DNA. They combine many of the properties of both RNA and DNA viruses. Only a small number of riboviruses are oncogenic and all belong to the genus Oncovirinae. These viruses are enveloped and have a single-stranded RNA molecule of mol. wt. 3 x 106.
The synthesis of DNA by RNA viruses, once regarded as heresy to the doctrine which stated that information flowed from DNA to RNA to protein, now has a place of honor in molecular biology. Viruses that do so are oncogenic riboviruses. Earlier studies showed that DNA played a critical role in the multiplication of RNA tumor viruses and in their ability to transform cells. Infection and transformation by these viruses can be prevented by inhibitors of DNA synthesis added the first 8-12 hr. after exposure of cells to the virus.
Also, the formation of virions is sensitive to actinomycin D suggesting a requirement for DNA-dependent RNA synthesis. These data led Howard Temin of the University of Wisconsin to propose his provirus theory which postulated the transfer of the information of the infecting RNA to a DNA copy which then serves as a template for the synthesis of viral RNA. David Baltimore of the Massachusetts Institute of Technology, and Temin in 1970 reported the presence of an RNA-dependent DNA polymerase or reverse transcriptase, under the influence of which DNA was synthesized from an RNA template.
Viral RNA is converted into linear double-stranded DNA in the cytoplasm. This is DNA provirus. This provirus is transported to the nucleus and there integrated with host DNA (integration may not be required in some cases, and in such cases also retrovirus multiplication occurs). mRNAs are usually synthesized and transported to the cell cytoplasm. (BSc 2nd Year Microbiology Viruses Notes Study Material)
Representatives of most major families of deoxyviruses are associated with cancer. Amongst animal systems, the best understood in molecular terms and the smallest in genome size are the papovaviruses, polyomavirus, and simian virus 40.
Several papovaviruses cause warts (papillomas) in men and domestic animals. One causes no observable disease in laboratory mice but when artificially inoculated into infant rodents causes a wide variety of tumors, hence its name poly-oma. Another, simian virus 40 (SV 40), came to light during the preparation of the poliovirus vaccine in monolayers of monkey kidney cells. SV40 produces vacuoles in infected cells and is sometimes called a vacuolated agent. The name of this group papovavirus is thus derived from the initials of its chief members: PA for papilloma, PO for polyoma, and VA for a vacuolating agent. (BSc 2nd Year Microbiology Viruses Notes Study Material)
Cellular proto-oncogenes. Cellular proto-oncogenes are involved in oncogenesis by DNA tumor viruses as well as retroviruses. Originally they were discovered by their homology to retroviral oncogenes but the discovery that they were oncogenic in their own right established the cellular proto-oncogene theory. How can c-once be at one and the same time ‘normal’ genes and oncogenes? There may be four possibilities:
(i) their expression be enhanced,
(ii) be amplified,
(iii) be relocated,
(iv) suffer point mutations.
There is enhanced expression, as through the insertion of viral LTRs (long terminal repeat sequences) in retroviruses. Gene amplification is seen in many tumor cell lines in culture. (BSc 2nd Year Microbiology Viruses Notes Study Material)

Relocation of the cellular proto-oncogene c -myc is seen as one of the later stages in the transformation of B lymphocytes by EBV. Point mutation of a c-onc endows it with malignant properties. To date mutated c -ras genes have been identified in 70% of human tumors investigated. (BSc 2nd Year Microbiology Viruses Notes Study Material)
How viruses trigger cells to become tumor cells has been the subject of intensive research in recent years. Most studies have centered on the oncogene theory, first proposed by Robert Huebner and George Todaro in 1969. This theory suggests that the genes transform a cell, and the oncogenes reside in the chromosomal DNA of a normal cell. When a genetic change occurs, such as by the introduction of a virus, the oncogenes transform the cells into cancer cells. (BSc 2nd Year Microbiology Viruses Notes Study Material)
In the modern view of cancer researchers, the oncogenes are presumed to exist as genes called proto-oncogenes. These may have important functions in the normal regulation of cell growth and development, perhaps by turning on at specific stages of life and turning off later.
In 1985, the proto-oncogenes were linked to the production of cyclic adenosine monophosphate (CAMP). An organic substance is central to the regulation of many physiological processes. Proto-oncogenes have been demonstrated in diverse forms of life from yeasts to humans. Oncologists believe that chromosomal alterations by carcinogens convert proto-oncogenes into oncogenes that transform a cell.
The major carcinogens are radiation, some chemicals, and recently linked viruses. Viruses in lysogeny may transform a cell. DNA of a virus may attach to chromosomes like a prophage. However, in RNA viruses, an enzyme called reverse transcriptase synthesizes DNA from RNA. This DNA then binds to the cell chromosome. (BSc 2nd Year Microbiology Viruses Notes Study Material)
One mechanism of transformation has been studied in retroviruses. These viruses carry oncogenes which are inserted into a host cell’s chromosome along with the newly synthesized viral DNA. This process is like transduction. Once on the host cell’s chromosome, the oncogenes apparently switch on the cell’s proto-oncogenes and activate them into oncogenes that transform the cell.
In 1980, Robert Gallo and his co-workers at National Cancer Institute identified a retrovirus as the cause of cancer of the white blood cells called human T-cell leukemia. The virus believed to be the first linked to human cancer was named HTLV-I. In 1984 Gallo and his group identified another retrovirus, HTLV-III, as the agent of AIDS. (BSc 2nd Year Microbiology Viruses Notes Study Material)
Another mechanism of transformation has been studied with the virus of Burkitt’s lymphoma. A cancer of lymphoid connective tissues of the jaw. The viral genome perhaps inserts itself into a chromosome of B-lymphocytes. At present, over 20 oncogenes have been discovered in human cancers including those of the bladder, lung, and colon, and a number of oncogenes have been linked to the production of various proteins. (BSc 2nd Year Microbiology Viruses Notes Study Material)
Viroids
Viroids are distinct low molecular weight nucleic acids (1.1 to 1.3 x 105 or 100.000-125,000 daltons) that could be isolated from certain plants afflicted with specific diseases. Unlike viral nucleic acids, they are not encapsidated i.e. capsid is lacking. There is no virus-like nucleoprotein particle in infected tissue. Viroids constitute a novel class of subviral pathogens.
They are the smallest known agents of infectious disease. So far viroids are definitely known to exist only in higher plants. And to consist of RNA only with a unique, previously unknown structure. (BSc 2nd Year Microbiology Viruses Notes Study Material)
The first viroid was discovered in attempts to purify and characterize the causative agent of potato spindle tuber, a disease that for many years had been assumed of viral etiology. In 1967 Diener and Raymer reported that the transmissible agent of this disease was a free RNA and that no viral nucleoprotein particles (virions) could be detected in the infected tissue.
By 1971, sedimentation and gel electrophoretic analysis had shown conclusively that the infectious RNA was far smaller than the smallest genomes of autonomously replicated viruses (Diener, 1971). No evidence could be obtained for the involvement of helper viruses in the replication of the RNA. Thus, in spite of its small size, the RNA appears to be replicated autonomously in susceptible cells.
Because of the basic differences between the potato spindle tuber disease agent and conventional viruses Diener (1971) introduced the term viroid to denote it (potato spindle tuber agent) and agents with similar properties. Since then much attention has been paid to viroids and viroid diseases. A dozen diseases of higher plants are now known or believed to be caused by viroids. These are as follows:
(1) Potato spindle tuber (Diener, 1971).
(2) Citrus exocortis (Sanger, 1972; Semancik and Vanderwoude, 1972).
(3) Chrysanthemum stunt (Diener and Lawson, 1973).
(4) Chrysanthemum chlorotic mottle (Romaine and Horst, 1975).
(5) Cucumber pale fruit (Sanger et al., 1976; Van Dorst and Peters, 1974),
(6) Hope stunt (Sasaki and Shikata, 1977).
(7) Tomato planta macho (Galindo et al., 1980).
Some plant diseases suspected to be viroid diseases due to viroid-like RNAs are as follows:
(1) Coconut candangcadang (Randles, 1975).
(2) Avocado sunblotch (Mohamed and Thomas, 1980; Palukaitis et al., 1979).
(3) In plants of Columnea erythrophae, symptoms are like those of potato spindle tuber viroid. But the structure of the causative agent is different (Owens et al., 1978). (BSc 2nd Year Microbiology Viruses Notes Study Material)
It has been shown in studies made so far on different viroids that infection of the hosts occurs in a persistent manner i.e. no recovery occurs and viroids could be isolated from infected plants as long as the plants live. Transmission is mechanical. The symptoms of host plants are almost similar to those of viruses. Similarly, cross-protection also operates in viroid diseases.
Structure of viroids
Viroids are single-stranded, covalently closed circular as well as linear RNA molecules that occur because of extensive regions of intramolecular complementarity, in the form of collapsed circles and hairpin structures with the appearance in the electron micrographs of double-stranded molecules. Viroid RNA molecules are so small that the largest one so far described (CEV-citrus exocytic viroid) is only 371 nucleotides long-about one-tenth of the size of the smallest RNA virus. The complete primary sequence of potato spindle tuber viroid (PSTV) has been determined. A model for the secondary structure has been proposed by Gross et al. (1978).
According to this model: “viroids exist in their native configuration as extended rod-like structures characterized by a series of double helical sections and internal loops. Thus the rigid rod-like structure of the native viroid is based on a defective rather than a homogenous RNA helix.”
From a plant physiological viewpoint, viroids may be regarded as abnormal products of plant metabolism. Detailed viroid structure is given in the review by Gross and Riesner (1980). Other extensive reviews on the subject are by Diener (1977, 1979 a,b) and Diener and Haddi (1977). (BSc 2nd Year Microbiology Viruses Notes Study Material)
BSc 2nd Year Microbiology Viruses Notes Study Material
BSc 2nd Year Sample Model Practice Mock Test Question Answer Papers