BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material
BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material: BSc is a three-year program in most universities. Some of the universities also offer BSc Honours. After getting enrolled for BSc, there are certain things you require the most to get better grades/marks in BSc. Out of those, there are BSc Study Material, BSc Sample Model Practice Mock Question Answer Papers along with BSc Previous Year Papers. At gurujistudy.com you can easily get all these study materials and notes for free. Here in this post, we are happy to provide you with BSc 2nd Year Zoology Balanoglossus Notes Study Material.

BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material
Balanoglossus: A Tongue Worm
Hemichordata (Gr. hemi, half; chorde, cord), till recently treated as a subphylum of the phylum Chordata, is now regarded to be an independent phylum of invertebrates close to Echinodermata. However, for convenience of the comparative study of the protochordates, the Hemichordata is being retained here as a chordate subphylum. It includes a small group of soft, vermiform, marine and primitive chordates, commonly called the ‘acorn worms’ or ‘tongue worms’. The most familiar hemichordate genus is Balanoglossus which belongs to the class Enteropneusta. Other closely related genera are Saccoglossus (= Dolichoglossus), Glossobalanus, Ptychodera, Spengelia, etc.
Derivation of Name
Balanoglossus clavigerous was recorded and named by Delle Chiaje in 1829. Its generic name was derived from two Greek words, balanos and glossus. The term balanos means an ‘acorn’ (fruit of oak) and refers to the proboscis projecting from the collar looking like an acorn-nut, hence the common name “acorn worm “. The term glossus means ‘tongue’ and refers to the shape of its proboscis, collar and genital wings bearing a close resemblance to an ox tongue, hence the common name ‘tongue worm’. The local fishermen call Balanoglossus by the name ‘ox tongue’.
Geographical Distribution
Balanoglossus, like all other hemichordates, is a marine animal having worldwide distribution. About 20 species occur all over the world, especially in the tropical and subtropical seas. Some species are B. australiensis (Australia), B. carnosus (Indo-Pacific), B. misakiensis (Japan), B. jamaicensis (West Indies), B. gigas (Brazil), B. capensis (South Africa) and B. clavigerus (the Mediterranean and British isles).
Habits and Habitat
Habitat: Balanoglossus is a marine, tubicolous or burrowing hemichordate inhabiting shallow coastal waters of the intertidal zone, but a few occur in deeper water.
Burrow: The animal may conceal under stones or seaweeds or excavate its own burrow in the bottom sand or mud. B. clavigerus lives inside a U-shaped tube or burrow with two vertical limbs 50-75 cm deep and two openings 10-30 cm apart. In some species (Saccoglossus), the body of the animal and tube are twisted, whereas the anterior and posterior extremities remain straight. The anterior opening of the burrow is funnel-shaped and exposed. The anterior vertical limb may give outside branches each having its independent funnel-like opening. The posterior opening of the burrow is rounded and concealed below the spirally coiled faecal matter of the animal.
Protective device: The inner wall of the fragile tube is smoothly lined by sand particles cemented together into a tough cast with sticky mucus secreted by the skin mucous glands of the animal. This prevents the collapse of the burrow and protects the delicate animal from burial in loose sand. Another protective device is the secretion of a foul-smelling odour similar to iodoform. One species shows phosphorescence. (BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material)
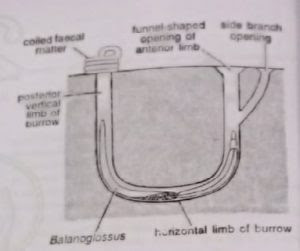
Movements: The worm is sluggish and little affected by external stimuli. It moves in its burrow by cilia covering its body surface. The most active part of the body is the proboscis. It elongates and shortens muscular activity and helps in burrowing. When the tide recedes, the tongue worm protrudes its anterior end out of the burrow to explore the surroundings, or its posterior end to cast out a faecal matter.
Feeding and breeding: It swallows sand or mud to obtain diatoms, protozoans, other microorganisms and organic detritus on which it feeds. Sexes are separate. Males and females, living in separate tubes, shed their gametes in seawater where fertilization occurs. The life cycle includes a free-swimming planktonic ciliated larva, the tornaria. Tongue worm does not reproduce asexually, but their fragile body has considerable power of regeneration.
External Morphology
Shape, size and colouration: The body is soft, elongated, worn-like, cylindrical and bilaterally symmetrical. It measures 10 to 50 cm in length, according to species. B. gigas attains a length of 1.8 m (Sawaya, 1951) or 2.5 m (Spengel, 1893). The Colour is bright or drab with reddish or orange tints. The body is uniformly ciliated and without any exoskeleton or external appendages.
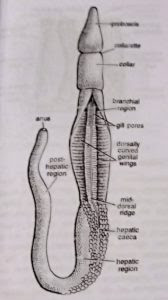
Division of body: The body is unsegmented but divisible into three distinct regions or parts: proboscis, collar and trunk
- Proboscis: The proboscis or protosome is the anteriormost part of the body. It is short, club-shaped or conical and circular in cross-section. It has a thick muscular wall and is hollow within. Its cavity or proboscis coelom communicates with the outside through a minute probosci’s pore situated mid-dorsally near its base.
In some species, the probosci’s pore ends blindly or there are two pores. Posteriorly, the proboscis narrows into a slender neck or probosci’s stalk which is attached to the collar. Below the stalk, the base of the proboscis bears a U-shaped ciliated epidermal depression called the preoral ciliary organ, which tests the quality of food and water entering the mouth. - Collar: The collar or mesosome is the middle, short and cylindrical part. Its flap-like or funnel-like anterior margin termed collarette completely surrounds and conceals the proboscis stalk and the posterior part of the proboscis. Ventrally, below the probosci’s stalk, the collarette or collar-nm encloses a permanently open wide aperture, the mouth. It opens into the buccal cavity inside the collar. The posterior end of the collar is well-demarcated from the trunk by a circular constriction.
The wall of the collar is thick, and highly muscular and encloses a cavity, the collar coelom. It opens outside through a pair of collar pores into the first pair of gill pouches. - Trunk: The trunk or metasome is the posterior and largest part of the body. It is rather fat and appears annulated due to circular constrictions on the surface. All along its length, the trunk bears a mid-dorsal and a mid-ventral ridge, each accommodating its corresponding nerve and blood vessel. The trunk is further differentiated into three regions: an anterior branchiogenital, a middle hepatic and a posterior post-hepatic, abdominal or caudal region.
(a) Branchiogenital region: The anterior or branchiogenital region of the trunk is marked by a pair of lateral, thin, flat and longitudinal flaps, the genital wings, containing the gonads. The gonopores are microscopic and cannot be seen by the unaided eyes. The anterior half of branchiogenital region bears two longitudinal rows of small branchial apertures or gill pores. One row of gill pores is mounted on a prominent longitudinal ridge on each side of the mid-dorsal ridge. The number of gill pores increases with the age of the animal.
The two genital wings can be curved to meet mid-dorsally so as to conceal the gill pores. In some species, a posterior prolongation of the collar, called the operculum, may cover the anteriormost gill pores.
(b) Hepatic region: The middle or hepatic region of the trunk is somewhat smaller than the genital region It is greenish in colour and its dorsal surface is marked by the presence of numerous irregular intestinal sacculations of hepatic caeca.
(c) Posthepatic region: It is the posterior-most and the longest part of the trunk, also called the abdomen or the caudal region. It is more or less uniform in diameter but its posterior end tapers slightly and bears a terminal anus.
Body Wall
The body wall is composed of the epidermis, musculature and peritoneum.
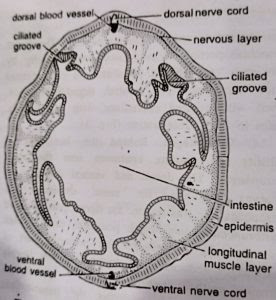
1. Epidermis: The outermost layer or epidermis consists of a single layer of mostly tall, slender, columnar and ciliated cells. Three kinds of gland cells secreting mucus are present. Goblet gland cells are flask-shaped with fine granules. Reticulate gland cells have mesh-like vacuolated cytoplasm. Mulberry or granular gland cells contain coarse granules and also secrete amylase.
The gland cells are more abundant in the collar region. Besides, neurosensory cells, which stain darker, are present in the epidermis of the proboscis and the anterior part of the collar. The dermis is absent. Below the epidermal cells is a thick nervous layer consisting of a network of nerve cells and nerve fibres. Below the nervous layer is a thick basement membrane which supports the epidermis and serves for the attachment of the underlying musculature. (BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material
2. Musculature: The muscles are smooth, weak and mostly longitudinal. The proboscis and the anterior end of the collar (collarette) have an outer layer of circular muscle fibres and an inner layer of longitudinal muscle fibres. In the trunk region, only longitudinal muscle fibres are present. (BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material)
3. Peritoneum: The coelom is lined by the parietal coelomic epithelium or peritoneum which covers the inner surface of the body wall musculature.
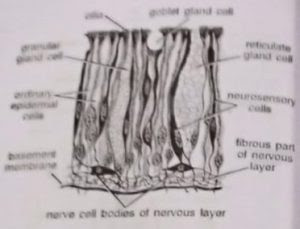
Functions of body wall: (i) The body wall shields the soft internal organs from mechanical injuries. (ii) Mucus produced by epidermal gland cells adheres sand particles for lining the burrow in which the worm lives. (iii) Foul smell of mucus is protective. (iv) Neuro-sensory cells serve to receive external stimuli. (v) Musculature helps in body movements.
Coelom
The coelomic cavities of the proboscis, collar and trunk are completely separated from one another. However, in the adult, the original coelom is greatly obliterated by connective tissue and muscle fibres derived from coelomic epithelium which becomes inconspicuous and irregular. These include one cavity in the proboscis, two in the collar and two in the trunk.
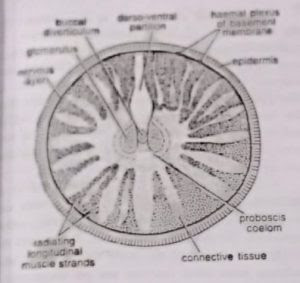
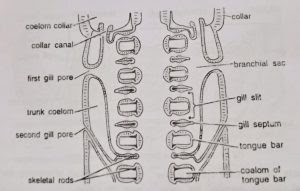
- Proboscis coelom: The unpaired proboscis coelom or protocoel is greatly obliterated by connective tissue and muscle strands except for a small central space which is occupied by the probosci’s complex. The latter includes the buccal diverticulum central sinus, heart vesicle and glomerulus. The proboscis coelom communicates with the outside through a proboscis canal and a proboscis pore situated mid-dorsally at the base of the probosci’s stalk.
- Collar coelom: The collar coelom or mesocoel is represented by two narrow lateral cavities, one on each side between the collar wall and the buccal tube. The two cavities are partitioned by incomplete mid-dorsal and mid-ventral mesenteries. The collar coelom does not communicate with the proboscis coelom, but, posteriorly, each cavity opens into the first-gill sac of its side by a collar canal and a collar pore. The collar coelom is greatly obliterated by the collar musculature and connective tissue.
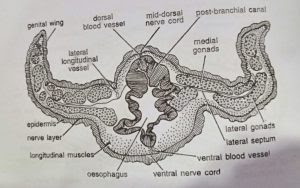
- Trunk coelom: The trunk coelom or metacoel is represented by a pair of closed cavities between the body wall and the gut wall. The two cavities are separated by an incomplete dorsal and a complete ventral mesentery. In the branchiogenital region, each cavity is further divided by a lateral septum into a dorsal-lateral and ventro-lateral compartment. The trunk coelom is partitioned from the collar coelom by a collar-trunk septum. The trunk coelom is obliterated by the trunk musculature.

Coelomic fluid: The proboscis and collar coeloms communicate with the exterior and are largely filled with seawater which keeps them turgid. The trunk coelom is filled with a watery coelomic fluid containing amoeboid coelomocytes, each with a single large vacuole. The Coelomocytes originate from the coelomic epithelium. According to Spengel, they behave like leucocytes by secreting a membrane around any foreign body that may invade the animal. (BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material)
Endoskeleton
Balanoglossus has no definite endoskeleton of bone or cartilage. However, the following four stiff structures are present: (i) Buccal diverticulum, (ii) proboscis skeleton, (iii) branchial skeleton, and (iv) pygochord.
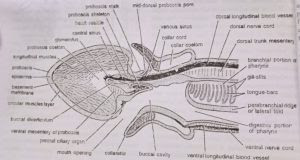
- Buccal diverticulum: From the roof of buccal cavity (lying inside collar region), a short stiff, thick-walled, hollow projection extends forwards through the proboscis stalk into the proboscis coelom. Its wall is composed of a single layer of tall, slender, vacuolated endodermal cells. For a long time it was considered a notochord (Bateson, 1885), or stomochord (Willey, 1899 and Dawydoff, 1948). Histoiogically as well as developmentally, it is quite different from the true notochord of other chordates, but resembles the wall of the buccal cavity. To most modem workers it is only a preoral extension of buccal cavity, therefore, Hyman preferred to use the noncommittal term buccal diverticulum for this tubular outgrowth of buccal cavity.
- Proboscis skeleton: The proboscis or nuchal skeleton is a Y-shaped chitinous structure formed by the thickening of the basement membrane. It consists of a broad, flat, roughly rectangular median plate produced ventrally into a keel and posteriorly into two diverging hors. The median plate lies below the buccal diverticulum in the proboscis stalk while the two horns extend backwards into the roof of the buccal cavity. The median plate remains embedded in a reticulum of stiff chondroid tissue which resembles vertebrate cartilage.
- Branchial skeleton: It is also formed by the thickening of the basement membrane. It consists of numerous M-shaped chitinous skeletal rods that lie in the wall of the pharynx and support the U-shaped gill slits that perforate it. The middle arm of the skeletal rod is thicker than the others and bifurcated at the free end which indicates that it is formed by the fusion of two arms of two adjacent ‘inverted U-shaped’ rods that join to form the M-shaped rod.
- Pygochord: In the post-hepatic region of the trunk, mid-ventrally between the intestine and body wall, develops a rod-like thickening called pygochord. Its cells are vacuolated. It supports the post-hepatic region of the body but probably also performs some other function not yet understood. (BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material)
Digestive System
[I] Alimentary canal
The alimentary canal is a complete and straight tube running between the mouth and anus. It is supported throughout its length by the dorsal and ventral mesenteries. Its wall is made up of ciliated epithelium covered externally by a basement membrane, but peculiarly, muscle layers are absent. Alimentary canal comprises of (i) mouth, (i) buccal cavity; (iii) pharynx, (iv) oesophagus, (v) intestine, and (vi) anus.
- Mouth: It is a wide and circular opening situated ventrally in a groove between the proboscis stalk and collarette. According to Knight-Jones (1952), it can be closed or opened and does not remain permanently open as previously supposed. It has two sets of muscle fibres, the radial fibres to open it and the concentric fibres to close it. The mouth leads into buccal cavity.
- Buccal cavity: The short buccal cavity occupies the collar region. Its epithelial wall contains glandular goblet cells. Anteriorly its dorsal wall forms a short, stiff and hollow buccal diverticulum: that projects into the proboscis coelom. Posteriorly it extends up to the collar-trunk septum behind which it continues into the pharynx.
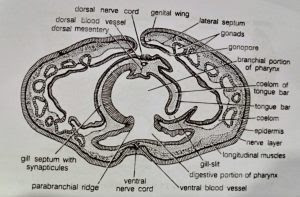
- Pharynx: It lies in the branchial region of the trunk. Externally its wall bears a longitudinal constriction along each lateral side. These lateral constrictions project into its lumen as ridges, called parabranchial ridges, consisting of tall columnar cells. These ridges incompletely divide the pharynx into a dorsal respiratory or branchial portion and a ventral digestive portion. The dorsal branchial portion is perforated dorso-laterally by two rows of U-shaped gill-slits, and is concerned with respiration. The ventral digestive portion, lined with ciliated epithelium with gland cells, helps in food concentration.
- Oesophagus: Behind the last pair of gill slits the pharynx continues into the short oesophagus. The dorsal and ventral divisions of pharynx continue for some distance into oesophagus. In this region, the dorsal part is called postbranchial canal which possesses thick, folded and glandular epithelium. The posterior part of oesophagus reduces in diameter and has deeply furrowed epithelium.
- Intestine: It occupies the hepatic and posthepatic regions of trunk. The hepatic region of the intestine is highly vascular. Its epithelial cells are dark green or dark brown, and its dorsal wall forms numerous sacculations called hepatic caeca. The intestinal wall lies in close contact with the body wall, so that the intestinal sacculations correspond with those of the body wall. The post-hepatic region of intestine is connected with the ventral body wall by the pygochord described earlier. It is a simple and straight tube bearing a pair of dorso-lateral grooves lined by tall epithelial cells with long cilia.
- Anus: Posteriorly, the intestine opens to the exterior by a terminal circular aperture, the anus, at the tip of the trunk. It is often surrounded by a sphincter muscle.
[II] Food, feeding and digestion
Balanoglossus is a ‘ciliary feeder’. Its food comprises of microscopic organisms and organic particles present in water and the bottom sand in which it makes its burrows. The lateral cilia lining the gill-slits set up a current of water which enters through the mouth, takes its course through the buccal cavity, pharynx, gill-slits and branchial sacs, and leaves through the gill pores. This is the respiratory-cum-food current.
Some food particles directly enter the mouth with this current while some come in contact with the proboscis and get entangled in the mucus that covers it. The mucus is secreted by the gland cells of the proboscis epithelium. Cilia covering the proboscis direct the mucous string, containing food particles, towards the pre-oral ciliary organ at the base of the proboscis. From here the mucous string is passed back into the mouth by the action of the proboscis cilia, aided by the main water current entering the mouth. Organic particles present in the sand are ingested directly along with the latter at the time of burrowing.
The U-shaped pre-oral ciliary organ, at the base of proboscis stalk, tests the quality of food and water entering the mouth. Undesirable substances are prevented from entering the mouth by the ventral part of the collarette, which does so by covering the mouth. Thus, the rejected particles, instead of entering the mouth, pass back over the collar.
Backward movement of food through the alimentary canal is maintained by the cilia lining its walls. In the pharynx, the food moves through the ventral digestive portion Digestion is brought about by enzymes secreted by gland cells of the pharynx, oesophagus and hepatic region of the intestine. The exact process of digestion in Balanoglossus is not known. Undigested substances, along with sand and silt, pass out through the anus as ‘castings’. (BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material)
Respiratory System
The respiratory apparatus of Balanoglossus comprises: (i) the branchial portion of pharynx bearing gill-slits, and (ii) the branchial sacs that open out through gill-pores.
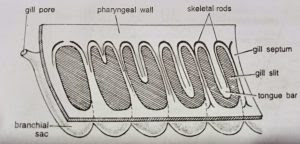
- Branchial pharynx: As already described, two lateral longitudinal parabranchial ridges divide the pharyngeal cavity into a ventral digestive portion and a dorsal respiratory or branchial portion. Dorso-laterally, on each side, the branchial portion is perforated by a longitudinal series of numerous U-shaped openings, the gill-slits. Their number varies and increases as the animal grows older.
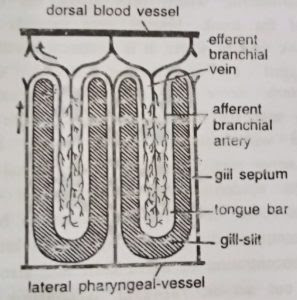
To start development a gill-slit is a broad oval slit. Later, a hollow projection of dorsal pharyngeal wall, called tongue bar, grows into the slit making it U-shaped. The hollow tongue bars enclose coelomic cavity and do not touch the ventral side of gill-slits. The portions of the pharyngeal wall between two adjacent gill-slits are termed gill septa. They are solid, without enclosing coelom. A tongue bar is connected with adjacent gill septa by short transverse or horizontal connections, the synapticula. The development and arrangement of gill-slits is identical with that found in Branchiostoma. (BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material)
As described earlier, the tongue bars and septa are supported internally by a chitinous skeleton forming M-shaped rods. A septum contains the middle arm of an M-rod which is bifurcated at its lower free end. The lateral arms of M-rod lie in adjacent tongue bars so that each bar contains two arms of two adjacent skeletal rods. The gill-slits are richly lined by cilia, called lateral cilia.
- Branchial sacs: Gill-slits do not open directly to outside. Each gill-slit opens into a gill pouch called branchial sac, which lies between the bodywall and the pharynx. Each branchial sac in turn opens to the exterior by a small, independent gill pore. However, in one species (B. misakiensis) the first four pouches become united to open by a common gill pore to outside. The collar coelom also communicates with the common branchial sac of its side through a collar canal. The gill-pores are visible externally in two longitudinal rows one on each side of the mid-dorsal ridge in the branchiogenital region of the trunk.
Mechanism of respiration: The lateral cilia lining the gill-slits set up a food-cum-respiratory current of water. It enters the pharynx through mouth, then passes through gill-slits into the branchial sacs and finally leaves through the gill pores. The tongue bars are richly vascular and participate in gaseous exchange. The blood of their capillary networks takes up the oxygen dissolved in water and returns carbon dioxide to it. (BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material)
Blood Vascular System
The blood vascular system of Balanoglossus is of the open or lacunar type. It consists of: (i) a colourless blood, (ii) a central sinus and a heart vesicle, (iii) distributing vessels or arteries and sinuses, and (iv) collecting vessels or veins.
- Blood: The blood is a colourless fluid containing few white corpuscles which are possibly detached endothelial cells. A respiratory pigment is probably absent. The functioning of the circulatory system is not properly understood.
- Central sinus and heart vesicle: The central sinus is a small elongated non-contractile sinus situated in the proboscis just above the buccal diverticulum. Just above it is a closed triangular cardiac sac or heart vesicle. Its ventral wall is muscular and contracts rythmically thereby producing pulsations in the central sinus that help in the circulation of blood. The central sinus receives blood from collecting vessels that open into its posterior end. Anteriorly, it pumps its blood into several afferent vessels which form a plexus in the proboscis gland or glomerulus, lying in front of it. In glomerulus the blood gets rid of its excretory wastes.
- Distributing vessels or arteries: Blood from the glomerulus is carried away by four arteries. Of these two arteries, a mid-dorsal proboscis artery and a mid-ventral proboscis artery, supply the proboscis. The other two, called efferent glomerular arteries, run backward along the two sides of buccal diverticulum, encircle the buccal tube as peribuccal vessels (which are actually of the nature of blood plexuses) and unite in a single longitudinal ventral vessel that runs up to the posterior end of the body through the ventral mesentery.
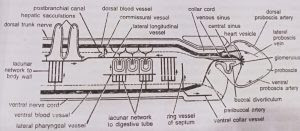
On its way, the ventral vessel gives out a ventral collar vessel to the collar, a ring vessel to the collar-trunk septum and an the afferent branchial artery to each gill septum in which it bifurcates to supply two adjacent tongue bars. All these branches break up into a system of sinuses in their respective structures. All along its length, the ventral vessel also supplies the body wall and gut wall by an elaborate network of sinuses. The ventral vessel has muscular contractile walls and the blood in it flows backwards.
- Collecting vessels or veins: Blood from body wall, gut wall and branchial apparatus (efferent branchial vessels) is collected by a single median dorsal vessel which runs through the dorsal mesentery, from posterior end upto the collar. It has muscular and contractile walls and blood flows in it forwards. At the anterior end of collar, the dorsal vessel dilates a little to form a venous sinus. The latter receives a lateral proboscis vein from each side of proboscis and then opens into the central sinus.
Excretory System
The excretory organ is glomerulus or proboscis gland lying in front of the central sinus and projecting into the proboscis coelom. It is made up of several blind tubular projections formed by the peritoneum covering the buccal diverticulum, central sinus and heart vesicle. The tubular projections contain blood confluent with that of central sinus. The excretory peritoneal cells of glomerulus contain yellow or brown granules, probably of excretory substances. From glomerulus the excretory substances pass on into the proboscis coelom and finally to the exterior through the proboscis pore.
Nervous System
The nervous system is of primitive type resembling that of coelenterates and echinoderms. Throughout the body a plexus or layer of nerve cells and nerve fibres lies just below the epidermis. Nerve fibres are traversed by the filamentous basal portions of epidermal cells, and form synapses with the processes of nerve cells. Nervous layer is thickened along definite strands to form two main nerve cords, one mid-dorsal and other mid-ventral, which runs along the entire length of the trunk. Ventral cord extends upto collar-trunk septum where it is connected with the dorsal cord by a circular strand, called circumenteric nerve ring.
Dorsal cord extends anteriorly upto the base of proboscis where it is connected with another circular strand called anterior nerve ring. In collar region, dorsal cord leaves the epidermis and traverses the collar coelom as collar cord. It is supposed to be the nervous centre of the animal. But unlike brain, it has no concentration of nerve cells and also does not give out nerves. However, it contains some giant nerve cells which help in transportation of impulses over the body and in reflexes. The collar cord contains a cavity called neurocoel.
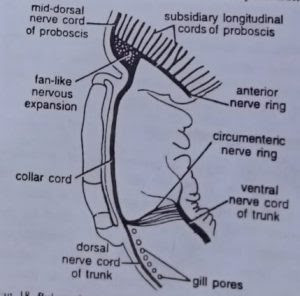
Sense organs of Balanoglossus are simple and comprise: (i) neuro-sensory cells in the epidermis of proboscis and anterior part of the collar, sensitive to touch and light; and (ii) preoral ciliary organ situated ventrally at the base of proboscis, which is a chemoreceptor.
Reproductive System
Asexual reproduction: Asexual reproduction is rare in enteropneusts. Gilchrist (1923) has described it in one species, B. capensis, which lacks hepatic caeca. During summer the young worms (juveniles) cut off small pieces from the tail end each regenerating into a complete sexual adult in winter.
Regeneration: Balanoglossus shows great power of regeneration. Proboscis, collar and isolated pieces from trunk can regenerate the lost parts of the body completely.
Sexual reproduction: Sexes are separate. Males and females cannot be identified externally except for the difference in colour of the ripe gonads in the living specimens. Testes of male and ovaries of female are similar. They are sac-like bodies occurring in several longitudinal rows in the genital wings, on either side of the alimentary canal. Each gonad has a narrow neck or ductule that opens out through a gonopore. All gonopores are situated outer to the gill pores. Ova are small with poor yolk content.
Affinities and Systematic Position of Balanoglossus (Hemichordata)
The group Enteropneusta, to which Balanoglossus belongs, was established by Gegenbaur in 1870. Bateson (1885) proposed the name Hemichordata in place of Enteropneusta. Since then, due to their peculiar anatomical organisation and embryology, the Hemichordata (or Balanoglossus) have been considered closer to the Chordata as well as most non-chordate phyla by different workers from time to time. Some of these views regarding the phylogenetic relationship (affinities) and taxonomic position of the hemichordates are summarized below.
[I] Affinities with Chordata
Some earlier workers, such as William Bateson (1885), proposed closer affinities between Hemichordata and Chordata. Their resemblance was based on the presence of the three fundamental chordate characteristics in Hemichordata, that is, (i) a notochord, (ii) a dorsal hollow nerve cord and (iii) the pharyngeal gill-slits (pharyngotremy).
Affinities with Urochordata: Hemichordates are nearest to Urochordata, as they exhibit many close resemblances with them. The structure and function of pharynx and branchial apparatus in hemichordates are similar to those of urochordates. Also, the development of the central part of nervous system is quite similar in both.
Affinities with Cephalochordata; Besides similarities in the structure and function of the branchial apparatus, the hemichordates also show similarity with cephalochordates in the arrangement of coelomic sacs and in development.
Due to these similarities Hemichordata had been considered as a subphylum of the phylum Chordata till recently, representing its lowest group, and probably having a common ancestry.
Objection: However, the hemichordates are no longer included under chordates because they do not possess chordate characters in a typical condition. The main objections are:
(1) A true notochord does not occur in hemichordates. Unlike that of the chordates, the so-called ‘notochord’ is very short, confined to proboscis and without any supporting function. It is ventral to the main (dorsal) blood vessel and not covered by sheaths. Instead of being solid and made of vacuolated cells, it is hollow and lined by epithelial cells. It does not originate from the roof of larval archenteron but as a forward hollow projection of the foregut. Instead of being called notochord (Bateson, 1885) it is now termed the stomochord (Willy. 1899). Hyman prefers to name it as buccal diverticulum.
(2) The nervous system is distinctly of the invertebrate type being intra-epidermal in position and having a ventral nerve cord and a circumenteric nerve ring which are absent in chordates. In Balanoglossus, the dorsal tubular nerve cord is confined to the collar region only.
(3) Gill-slits of Balanoglossus are numerous and dorsal in position, whereas they are 5 to 7 and lateral in higher chordates.
Other differences: The hemichordates further differ from the chordates in:
(1) Lacking metameric segmentation, cephalization, paired appendages, postanal tail, exoskeleton, living endoskeleton, dermis, liver, haemoglobin, red blood corpuscles, etc.
(2) Having peculiar division of body and coelom (into proboscis, collar and trunk), single-layered ciliated epidermis, hepatic caeca, dorsal heart, open neurocoel, colourless blood, numerous gonads, etc.
[II] Affinities with Rhynchocephalia
Feeding and burrowing habits are similar in Balanoglossus and Nemertinea. Body in both is elongated, vermiform, without external metamerism, with terminal anus, with smooth skin containing unicellular glands and ectodermal nerve plexus, and having metamerically arranged simple gonads. But Nemertinea differ in lacking a dorsal nerve cord and in having lateral nerve cords and a protrusible proboscis.
[III] Affinities with Phoronida
Some zoologists like A.T. Masterman (1897) advocated relationship of Balanoglossus with Phoronis on the following grounds:
(1) Similar nature of epidermal nervous system.
(2) The paired gastric diverticula of Phoronis, like the buccal diverticulum of Balanoglossus, forming so-called notochord.
(3) Actinotroch larva of Phoronis has several enteropneust features of tornaria such as similar disposition of coelom, anus surrounded by a ciliary ring presence of a proboscis pore and a sensory apical plate with cilia and eye spots.
(4) Both have great power of regeneration.
Objection: But the chordate features of Balanoglossus like pharyngeal gills are absent in Phoronis which also differs in having paired metanephridia. Moreover, Selys-Long Champ’s (1940) account of development of Phoronis does not coroborate Masterman’s observations, so that relationships of these two groups are rejected. (BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material)
[IV] Affinities with Pogonophora
Marcus (1958) tried to relate Hemichordata with Pogonophora due to following similarities:
(1) Enterocoelous formation of coelom.
(2) Body and coelom divided into three regions.
(3) Mesosome and metasome separated by a septum.
(4) Nervous system intra-epidermal.
(5) Pericardial sac in some pogonophores.
(6) Gonads found in trunk.
Objection: But pogonophores differ in having protocoelic nephridial coelomoducts and lacking an alimentary canal. Moreover, nervous system is concentrated in protosome in Pogonophora, but in mesosome in Hemichordata.
[V] Affinities with Annelida
Spengel (1893) first suggested affinities of Annelida and Hemichordata as follows:
(1) Body vermiform and coelomate.
(2) Burrowing habit, tubicolous life and ingesting mud which is passed out as castings through anus.
(3) Collar of Balanoglossus similar to clitellum of earthworm.
(4) Proboscis and prostomium similar and preoral.
(5) Similar arrangement of blood vessels with blood flowing anteriorly in dorsal vessel and posteriorly in ventral vessel.
(6) Dorsal position of heart.
(7) Tornaria larva of Balanoglossus shows several structural resemblances with the trochophore larva of Annelida in being pelagic, ciliated with apical plate, eye spots, sensory cilia and well developed alimentary canal with similar parts.
Objection: However, the two groups show striking differences as follows:
(1) Annelids do not nave pharyngeal gill-slits, stomochord or buccal diverticulum and dorsal tubular nerve cord found in Balanoglossus.
(2) Balanoglossus does not have double and solid ventral nerve cords and nephridia found in annelids.
(3) In tornaria larva of Balanoglossus, preoral or proboscis coelom is present, nephridia are absent and blastopore becomes anus of the adult (Deuterostomia). In trochophore larva of annelids, preoral coelom is absent, nephridia present and the blastopore becomes the mouth (Proterostomia).
Thus, compared to their great fundamental differences, the similarities of the two groups are only superficial and quite insignificant indicating probably a convergent evolution due to similar pres habits and habitat.
[VI] Affinities with Echinodermata
Adult resemblances: Adult hemichordates and echinoderms are structurally quite different and it is difficult to suspect any phylogenetic relationship the between them. They show few resemblances such com as:
(1) Enterocoelic origin of coelom and its division into three successive parts filled with sea water to serve a hydraulic mechanism.
(2) Heart vesicle and glomerulus of enteropneusts are considered homologous to the dorsal sac and axial gland of echinoderms. Both the structures are related and combine vascular and excretory functions.
(3) Nervous system is poorly developed and forms epidermal nerve plexus.
(4) Proteins and phosphagens present in hemichordates closely resemble those of echinoderms.
(5) Common habits and ecological niches and remarkable power of regeneration.
Larval resemblances: The two groups show a strong affinity on embryological ground as the tornaria larva of Balanoglossus has a striking structural similarity with an echinoderm larva, in particular the bipinnaria larva of asteroids. In fact, the tornaria was regarded an echinoderm larva for a long time by Johannes Muller (1850), Krohn (1854), Agassiz (1864), etc., till Metschnikoff (1870) proved it to be an enteropneust larva. The larvae of the two groups possess the following common features:
(1) Small, pelagic, transparent and oval.
(2) Identical ciliated bands taking up a similar twisted course.
(3) Enterocoelic origin and similar development of coelom.
(4) Proboscis coelom opening to outside by proboscis pore of tornaria comparable to hydrocoel of echinoderm dipleurula.
(5) Bastopore becomes the anus (Deuterostomia) and digestive tract is complete with mouth, anus and same parts.
Objection: However, the tornaria larva shows presence of apical plate with sensory hairs and eye spots and telotroch which are absent in echinoderm larvae. The protocoel is single in tornaria but paired in echinoderm larva. This raises doubts about the echinoderm affinities of hemichordates. Fell (1963) and others believe that their larval similarities are only because of convergent evolution due to same mode of habits and habitat. (BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material)
[VII) Systematic position and phylogeny
The peculiar anatomical organisation of Balanoglossus or hemichordates makes their systematic position uncertain and controversial. The earlier workers (Bateson, 1885) placed them as a subphylum under the phylum Chordata representing its lowest group. But the only chordate feature shown by them is the presence of pharyngeal gill slits.
Therefore, some recent workers like Van der Host (1939), Dawydoff (1948), Marcus (1958) and Hyman (1959) have chosen to remove hemichordates from the phylum Chordata and treat them as an independent invertebrate phylum. Since the group comprises only about 80 species, it is included in the category of a minor phylum. (BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material)
The name Hemichordata (Gr. hemi. half; chorde, cord) means they are ‘half’ or ‘part’ chordates, a fact that is undisputed. Therefore, in the present work, Hemichordata has been retained as a subphylum of Chordata to avoid any controversy.
BSc 2nd Year Zoology Balanoglossus A Tongue Worm Notes Study Material
BSc 1st Year Sample Model Practice Mock Test Question Answer Papers