Microbiology Resistance to Disease Notes Study Material
Microbiology Resistance to Disease Notes Study Material: BSc is a three-year program in most universities. Some of the universities also offer BSc Honours. Out of those, there is BSc Resistance to Disease Notes Study Material, BSc Sample Model Practice Mock Question Answer Papers & BSc Previous Year Papers. At gurujistudy.com you can easily get all these study materials and notes for free. Here in this post, we are happy to provide you with BSc 2nd Year Microbiology Resistance to Disease Notes Study Material.

Microbiology Resistance to Disease Notes Study Material
The human body is in fact a closed container like a doughnut. If a parasite is to establish, it must first penetrate through the protective layer of this container. Intact skin and its extensions into the gastrointestinal, respiratory, and urinary tracts represent the first major form of resistance to infection and disease. This form of resistance is general or nonspecific since it exists in all humans and is present from the earliest time of life. (Microbiology Resistance to Disease Notes Study Material)
The non-specific resistance is also referred to as nonadaptive or constitutive resistance and it includes protective mechanisms offered by surface layers; non-specific antimicrobial like lysozyme in natural secretions and animal extracts and like phenols in plant extracts; phagocytosis: inflammation; interferons etc. These all structures act non-specifically although their activity may be enhanced during infection. This type of resistance thus protects against all parasites. (Microbiology Resistance to Disease Notes Study Material)
Another form of resistance is specific. The specific resistance or inducible resistance-immunology or adaptive responses develop only in response to the presence of a particular parasite and are directed solely at that parasite. These are specific for certain antigenic determinants and are amplified by contact with those determinants. (Microbiology Resistance to Disease Notes Study Material)
The human body thus responds to foreign intrusion by developing three-way strategies for resistance. (i) immediate destruction of parasite on surface layers by secretion of some chemicals (enzymes as lysozyme) or through an acidic medium, (ii) immediate immune responses, collectively referred to as hypersensitivity, and, (iii) long-term immune responses, which take some time (hours or days). The first way is non-specific, whereas the second and third are specific and relatively more complex defense lines that involve the participation of cellular and non-cellular components of the host. (Microbiology Resistance to Disease Notes Study Material)
Nonspecific Resistance
This form of resistance involves a wide range of factors. This depends upon the general well-being of the individual and the proper functioning of the body’s systems. Therefore, nutrition, age, fatigue, sex, and climate are the determinants of this form of resistance.
One form of nonspecific resistance is called species immunity, which implies that a disease affecting one species will not affect another. For instance, humans do not contract hog cholera, while hogs do not contract polio. Cattle plague is unknown in humans, while gonorrhea does not occur in cattle. Such immunities are believed to be based on physiological and anatomical differences. In chickens, a physiological difference lends resistance to anthrax. The normal temperature of chickens is 45°C, at which anthrax bacilli do not grow well. At 37°C, susceptibility increases. This phenomenon was first demonstrated by Pasteur in 1878.
Racial immunities are those that exist among various races and people of the world. Most racial communities are due to nonspecific factors related to a people’s way of life. Such immunities may reflect the evolution of resistant humans. For instance, black Africans affected by the genetic disease sickle cell anemia do not suffer from malaria, presumably because the parasite cannot penetrate distorted red blood cells. According to some such immunities exist because parasites have adapted to the body’s environment. Americans, for example, view measles as a mild disorder.
In addition to species and racial immunities, the body possesses a number of identifiable processes for nonspecific resistance. These are briefly presented below:
[I] Mechanical and chemical barriers
As indicated earlier in the chapter, the intact skin and the mucous membranes that extend into the body cavities are among the most important resistant factors. Unless these are penetrated, the disease can not establish. As physical barrier intact body surfaces represent the first line of defense against microorganisms, physically blocking the entry of pathogens into the body. The outer surface of the skin layer is composed of keratin, a protein not readily degraded by microbial enzymes.
Keratin resists the penetration of water and is thus a potent barrier to microbes. However, penetration is common in everyday life. A cut or abrasion, for example, allows staphylococci to enter the blood, and an arthropod bite permits many organisms to pass the skin barrier. Yellow fever viruses, malarial parasites, certain rickettsiae, and plague bacilli are some examples. Other means of nicks, war wounds, injections, etc. (Microbiology Resistance to Disease Notes Study Material)
Some features of the mucous membrane provide resistance to parasites. For instance, cells of the mucous membranes along the lining of the respiratory tract secrete mucus which traps heavy particles and microbes in the air. Cilia of other cells then move the particles along the membranes up to the throat, where they are swallowed. Stomach acid then destroys any microorganism.
Lactobacillus spp. The normal flora of the vagina produces various acids, that lower the pH thus enhancing the resistance in the vaginal tract. Antibiotic treatment eradicates these bacilli that encourage diseases like candidiasis and trichomoniasis. In the urinary tract, the slightly acidic pH of urine promotes resistance to parasites and the flow of urine flushes microbes away.
Stomach acid which has a pH of about 2.0 acts as a natural barrier to the gastrointestinal tract. Most microbes are killed in this environment, except some like typhoid and tubercle bacilli, protozoan cysts, and viruses of hepatitis A and polio. Bile from the gall bladder also inhibits microbes in the duodenum. Enzymes of the duodenum also digest large molecules of microorganisms.
A nonspecific chemical inhibitor is the enzyme, lysozyme, a protein described by Alexander Fleming (who later discovered penicillin) in the early 1920s. Lysozyme is present in human tears and salvia. It digests the peptidoglycan of the cell walls of Gram-positive bacteria. Some chemicals within the body bind iron, thereby withholding this essential growth element from pathogens. By limiting the amount of available iron, these compounds limit the growth of pathogens. Lactoferrin and transferrin are examples of such iron-binding chemicals. Lactoferrin is present in tears, semen breast mill bile, and nasopharyngeal, bronchial, cervical, and intestinal mucosa secretions.
Transferrin is found in serum and the intercellular spaces of various tissues and organs. Another nonspecific chemical inhibitor is interferon, a group of substances produced by body cells in response to the entry of a virus. Interferons trigger the production of inhibitory substances that “interfere” with viral reproduction. The various nonspecific resistance mechanisms are summarised in Table.

[II] Phagocytosis or Cellular defense
Shortly after the verification of the germ theory of the disease of Koch, a Russian, Elie Metchnikoff (native of Ukraine) made a chance discovery that clarified how living cells could protect themselves against microorganisms. Metchnikoff observed that motile cells in the larva of starfish gathered around a wooden splinter placed within the cell mass. He suggested that the motile cells actively sought out and engulfed foreign particles in the environment to provide resistance. His theory of phagocytosis, published in 1884 was received with skepticism since it appeared to conflict with the antitoxin theory.
However, in succeeding years most persons appreciated phagocytosis. Metchnikoff later became an associate of Pasteur and was co-recipient of the 1908 Nobel Prize in Physiology or Medicine. (Microbiology Resistance to Disease Notes Study Material)
Phagocytosis is considered a major form of nonspecific defense in the body. The cells involved are called phagocytes. They are the polymorphonuclear cells (PMNs) (also called polymorphs or neutrophils). Neutrophils are the most abundant phagocytic cells in the blood.
These are produced in the bone marrow and continuously circulate in blood and monocytes of the circulatory system, as well as cells of the reticuloendothelial system (RES). This system, also called the mononuclear phagocyte system, is a collection of monocyte-derived cells that leave the circulation and undergo modification in the tissues. They include Kupffer cells of the liver and macrophages of the spleen, bone marrow, lymph nodes, brain, and connective tissues. (Resistance to Disease Notes Study Material)
RES cells are larger than monocytes and have more lysosomes and a longer life span. Some phagocytes are called resting cells, because they are stationary, while others are wandering cells because they are actively motile.
Phagocytosis implies the ingestion of particles. Some authors use the term endocytosis to unify the concepts of both phagocytosis and pinocytosis (the step of taking dissolved material into the cell). However, it could not receive much support.
Phagocytosis begins with an invagination and pinching of the cell membrane to form a phagocytic vesicle or phagosome. The phagosome then fuses with a lysosome, an organelle that contributes digestive enzymes, lysozyme, and an acidic pH to the digestion process. Lysosomal substances also increase the permeability of capillaries which brings more phagocytes to the area.
The process is completed as waste materials are egested from the phagocyte. A chemical attraction called chemotaxis exists between parasite and phagocyte. This attraction is mediated by an unidentified substance released by the parasite. The interaction between parasite and phagocyte is also enhanced by the presence of antibodies. These protein molecules attach to parasites and thereby increase adherence to the phagocytic cell at a specific receptor site. (Microbiology Resistance to Disease Notes Study Material)
In other situations, components of the complement system bind the parasite to the phagocyte. Enhanced phagocytosis is called opsonization, and the antibodies or complement components that encourage it are termed opsonins, from Latin meaning “to prepare for”. Opsonins were first described by a British, Almroth Wright in 1903, who viewed them as serum proteins that bind to invading parasites, making them more susceptible to phagocytosis. At that time it was a way of unifying Metchnikoff’s phagocytic theory with the antitoxin theory. The view of Wright still holds, but the modern view of opsonins is somewhat different, to be seen later.
The serum sample may be analyzed by its opsonic index. A rough environmental surface as the wall of a blood vessel or a fibrin clot helps the phagocyte in trapping a particle. This is called surface phagocytosis. Unlike B- or T- lymphocytes the activities of macrophages are not confined to a specific antigen. However, macrophages have receptors for the Fc region of IgG molecules, and such ‘armed’ macrophages have antibody specificity conferred upon them. (Microbiology Resistance to Disease Notes Study Material)
[III] Inflammation
This is a non-specific defense response by the body to an injury in the tissue. It develops after a mechanical injury as an injury or blow to the skin, or from exposure to a chemical agent such as an acid or bee venom. It may also be due to a physical agent such as heat or UV radiation or to a living organism such as a parasite.
The irritant initiates a process that limits the extent of the injury. Dilation of blood vessels leads to increased capillary permeability, a flow of plasma into the tissue, and fluid accumulation at the site of irritation. Neutrophils adhere to the vessels close to the injury and migrate through the wall to begin phagocytosis of the irritant. Soon, macrophages replace the neutrophils.
The area shows the four characteristic signs of inflammation: (i) a red color from blood accumulation – rubor; (ii) a warmth from the heat of blood – color; (iii) a swelling from the accumulation of fluid-tumor, and (iv) pain from an injury to the local nerves – dolor. A product of phagocytosis is a mixture of plasma, dead tissue cells, leukocytes, and dead bacteria known as pus. When this material becomes enclosed in a wall of fibrin through a clotting mechanism, a sac may form. This sac is the abscess or several abscesses or boil. When several abscesses accumulate, an enlarged structure called carbuncle results. (Microbiology Resistance to Disease Notes Study Material)
Inflammation and phagocytosis are thus related to one another. The object is to confine the irritant to the site of entry and to repair or replace the injured tissue.
[IV] Fever
Fever is an abnormally high body temperature that may provide nonspecific resistance to disease. There is a scientific belief that bacteria, viruses, etc. affect a region at the base of the brain – the hypothalamus, and stimulate it to raise the body temperature. Due to this cell metabolism increases and blood vessels constrict thus denying blood to the skin and keeping its heat within the body. The patient thus experiences cold skin and chills along with a fever. The increased metabolism in cells also encourages tissue repair as well as increases in the level of phagocytosis.
Specific Resistance (Immunology)
In this section, we shall examine specific defense mechanisms and the nature of the immune system. The study of specific resistance to disease arose at the end of the 1800s. No one was really sure how the body responded to disease. Medical microbiologists were, however, aware that certain proteins in the blood unite specifically with chemical compounds of microorganisms. The blood proteins were named Bence Jones proteins after Henry Bence Jones, a British physician who identified them in the urine of a patient in 1847.
In 1922 researchers from Johns Hopkins University showed that Bence Jones proteins (eventually known as antibodies) were unlike normal serum proteins. Until the 1950s, specific resistance to disease was virtually synonymous with immunity. By that time, vaccines for several diseases were available, and immunologists viewed their field as one associated with disease prevention. But the explosion of interest in biological science after World War II spilled over to immunology and soon it became apparent that specific resistance was a phenomenon with wider implications. (Microbiology Resistance to Disease Notes Study Material)
Immunity mechanisms thus developed into a distinct science of immunology in the 1950s with such phenomena as organ transplantation (it is associated with immunology because the rejection mechanism was a type of specific resistance), allergic reactions, cancer, and Rh disease of infants, all related with a specific resistance. In addition, both diagnostic laboratories (serological tests) and vaccine research were gaining much information from immunology. This led to the elucidation of the structure of antibodies in the 1960s and the foundations of immunology as a key discipline of our times. (Microbiology Resistance to Disease Notes Study Material)
Basic concepts of immunology
Three concepts critical to the study of immunology are:
1. Specificity. It refers to the host’s response to an individual agent.
2. Memory. It implies that once the body has responded to an agent, it will react vigorously during subsequent exposure. This is the reason why an episode of measles makes the person immune to future episodes.
3. Recognition of non-self. This means that the host will develop resistance to agents that are foreign to itself. However, this recognition sometimes fails to operate, such as in autoimmune diseases.
The above basic concepts revealed that there are two different forms of the immune response : (i) antibody-mediated immunity (also called humoral immunity), in which specific proteins, the antibodies are made when foreign antigens are detected. An antigen is any macromolecule that elicits the formation of an antibody and that can subsequently react with an antibody.
In this kind of response, plasma cells derived from certain white blood cells (B-lymphocytes) synthesize antibodies in response to the detection of a foreign macromolecule with antigenic properties. Antibodies, also called immunoglobulins are made in response to specific antigens and react with those antigens. (ii) cell-mediated immunity (also called cellular immunity) in which certain cells of the body acquire the ability to destroy other cells that are recognized as foreign or abnormal. This type of response depends on another class of lymphocyte cells – T- lymphocytes. These interact with “foreign” cells to destruct them.
There are also interactions between B- and T-lymphocytes and between these and other blood cells that establish an integrated defense network. We shall now examine these things in somewhat detail. (Microbiology Resistance to Disease Notes Study Material)
Antigens
Antigens are chemical substances that elicit a response by the body’s immune system. In the old literature, antigens were considered foreign substances that elicited antibody formation by the immune system. However, it is now understood that an antigen need not be foreign but may be a person’s own chemical compound and that the response may involve something other than antibodies. Thus, a broader definition became necessary.
There is a wide variety of substances that act as antigens. It may include proteins, bee venom, hemoglobin molecules, bacterial flagella, pili, and capsules. The key feature of the antigen is an area of activity called the determinant. This is a location on the antigen molecule, consisting of six to eight amino acids or carbohydrates with a total mol. wt. of about 750 daltons. The antigenic determinant site is somewhat analogous to the coenzyme portion of an enzyme molecule. A single antigen may have two or more antigenic determinants.
Proteins are the best antigens because they have the largest array of potential building blocks (amino acids). This diversity permits a huge variety of antigenic determinants. Some polysaccharides may also serve as antigens. Examples are capsular polysaccharides in pneumococci and the blood group antigens A and B which are polysaccharides complexed with protein. Most polysaccharides, however, are poor antigens because they lack chemical diversity and rapidly break down when introduced to the body. Nucleic acids are not good antigens for the same season. Lipids are generally non-antigenic. (Microbiology Resistance to Disease Notes Study Material)
Antigens usually have a mol. wt. of over 10,000 daltons. This large size implies that a variety of determinants exists in an antigen. It also ensures that the molecule will be phagocytized, a necessary first step in the immune response. Smaller molecules may be complex with tissue proteins or polysaccharides to form antigens. The small molecule, called a hapten, provides the key functional unit of the antigen. For example, a derivative of the penicillin molecule is the hapten that combines with protein to yield antigens in allergic reactions. Other haptens are found in poison ivy, poison oak, certain dyes, and cosmetics.
Specific immunologic tolerance
Normally a person’s immune response as the proteins and polysaccharides do not elicit an immune response as they are interpreted as “self”. It is believed that before birth the proteins and polysaccharides of the body cells contact and inactivate immune system cells that might later respond to them. The responsive cells are easily paralyzed in the fetal stage. Thus the individual becomes tolerant of “self” and later will respond only to antigens interpreted as “non-self”. This theory, termed specific tolerance, was first advanced by Frank MacFarlane Burnet and Peter Medawar, the 1960 Nobel Prize winners in Physiology or Medicine.
Antigens enter the body through a variety of portals including passage through mucous membranes of the respiratory tract and penetration through the skin by arthropod bites, injections, and wounds. They may also enter through the lining of the gastrointestinal tract despite the destructive effect of stomach acid and duodenal enzymes. Once within the body, the antigenic nature of a substance is determined by such factors as the dose and size of the antigen, the ability of phagocytes to degrade the antigen, and the proper functioning of the immune system.
Types of antigens
There are many kinds of antigens. Among the hundreds of thousands of different antigens some are classified into the following groups:
1. Autoantigens. These are a person’s own chemical substances that elicit an immune response when self-tolerance breaks down. This may result in an autoimmune disease, such as systemic lupus erythematosus.
2. Alloantigens. These are present in some but not all members of a given species. The A, B, and Rh antigens of the various blood groups in humans are typical examples. If an Rh-negative woman bears an Rh-positive child, a condition called hemolytic disease of the newborn (Rh disease) may develop. Other alloantigens are involved in transplantation.
3. Heterophile antigens. They are identical antigens found in apparently unrelated species of organisms. These are represented by the antigens found in most rickettsiae and in Proteus Ox 19 cells. Antibodies produced in response to these antigens are identified in the Weil-Felix test. Another heterophile antigen is found in sheep red blood cells and infectious mononucleosis viruses as well as in horses, whales, turtles, and humans with blood type A. (Microbiology Resistance to Disease Notes Study Material)
Origin of the Immune System
The immune system is a general term for complex series of cells, factors, and processes that provide a specific response to antigens. One effect of this response is to lend specific resistance to disease.
The immune system appears to originate in the fetus, about two months after conception. At that time, primordial cells, called stem cells, arise in the bone marrow and differentiate by a complex process into either of two types of cells: (i) erythropoietic cells which will become erythrocytes, and (ii) lymphopoiesis cells which will become the lymphocytes of the immune system. We shall follow the fate of the lymphopoietic cells. (Microbiology Resistance to Disease Notes Study Material)
Lymphopoietic cells follow either of two courses. (i) certain ones pass through a specialized organ of the thoracic cavity called the thymus. In humans, this flat bilobed organ lies just below the thyroid gland in the upper thorax, above the heart. It increases in size until the age of puberty and then disintegrates.
Within the thymus, lymphopoietic cells are modified to form thymus-dependent lymphocytes or T-lymphocytes. They are also called T-cells. After they emerge, the T-lymphocytes (or T-cells) move through the circulation and colonize the lymph nodes. spleen, tonsils, and other lymphoid tissues. (ii) other lymphopoietic cells pass through an organ that has been identified in the chick but not with certainty in humans. In the chick, the organ is a gland in the lower gastrointestinal called the bursa of Fabricius.
Lymphopoietic cells that pass through this gland are modified to form B-lymphocytes (B for bursa). They are also called B-cells. In humans, the analogous organ is thought to be the fetal liver or bone marrow. B-lymphocytes have chemical substances on their surfaces that distinguish them from T-lymphocytes. Like T-lymphocytes, the B-lymphocytes move through the circulation to colonize the lymph nodes and other lymphoid tissues. (Microbiology Resistance to Disease Notes Study Material)
The T-lymphocytes and B-lymphocytes occupy a central role in the immune system. Since they accumulate in lymph nodes lying along the lymphatic vessels, they encounter all antigens except those entering the cardiovascular system directly. The lymphocytes are also prominent in the tonsil and spleen, both of which are important in youth but have a diminished role in adults. B-lymphocytes and T-lymphocytes are segregated into discrete areas in the lymphoid tissues.
Usually, the T-lymphocyte is the smaller of the two cells. B-lymphocytes have a life span of about five to seven days, but T-lymphocytes may live for many months or years. About 65 to 80 percent of the circulating lymphocytes are T-lymphocytes, and most of the remainder are B-lymphocytes, with a small percentage of immature cells of either type.
The production of surface components of B-lymphocytes is under the control of genes called the immune response (Ir) genes. These genes also direct the synthesis of individual markers on different B-lymphocytes and the manufacture of antibodies in immune responses. In humans, Ir genes appear to occur on various chromosomes. (Microbiology Resistance to Disease Notes Study Material)
Operation of the Immune System
The immune process begins with the entry of antigens into the lymphatic or cardiovascular system as shown in Fig.
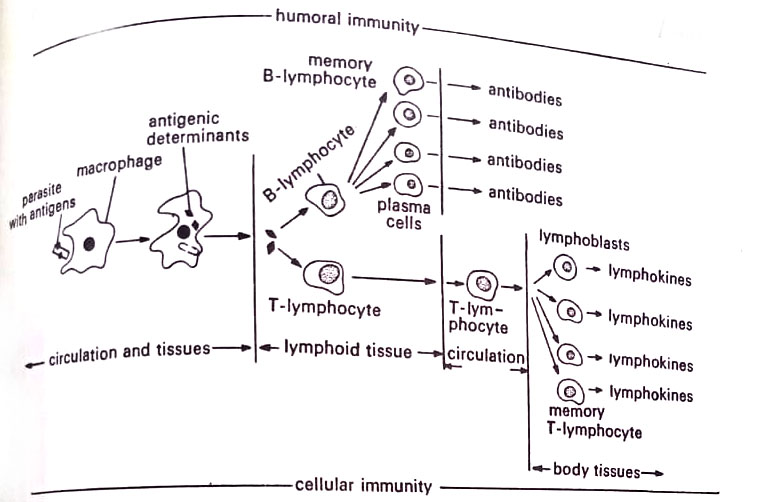
The antigens are phagocytized by macrophages, monocytes, or polymorphonuclear cells, and the major portion of the antigenic material is digested. However, the phagocytes preserve the antigenic determinants and transport them to the immune system in the lymphoid tissue. There is evidence that unprocessed antigens (unphagocytised antigens) are poor stimulators of the immune system.
In the lymphoid tissue, the macrophages present the antigenic determinants to T-lymphocytes and B-lymphocytes. The lymphocytes gather about the macrophages and the cells cling to one another. An interaction then takes place between antigenic determinants and specific receptor sites on lymphocytes. According to some immunologists, the antigenic determinants are released on the disintegration of the macrophages, while others present evidence of cytoplasmic channels between the receptor cells. A third contention is that the antigen is released in microscopic vacuoles. (Microbiology Resistance to Disease Notes Study Material)
At this point, the immunity process diverges depending upon whether T-lymphocytes or B-lymphocytes are stimulated. Two forms of immunity, cellular immunity, and humoral immunity, are possible. We shall discuss each process separately.
[I] Cellular immunity
The form of immunity arising from T-lymphocytes is termed cellular immunity because it happens on or near the body cells and is under the direct influence of lymphocytes. It is also referred to as cell-mediated immunity (influenced by cells), or tissue immunity because the immunity to antigens takes place within the tissues. The immunity develops as follows:
The antigens of many fungi and protozoa and selected viruses and bacteria stimulate the T-lymphocytes and sensitize them. Sensitized T-lymphocytes then enter the circulation and migrate to the site where the antigen was detected. The pool of lymphocytes increases as other T-lymphocytes, some from the circulation, are sensitized and drawn to the site by transfer factors from the original lymphocytes.