Ribosomes. They are seen in thin sections as relatively dense particles about 20 nm in diameter present as polysomes. Ribosomes can be easily obtained from ruptured cells by differential centrifugation on a sucrose gradient. They are made up of two sub-units of sedimentation constants 30S and 50S, which combine to give the characteristic prokaryotic 70S ribosome. Both components are made up of roughly equal amounts of RNA and protein. The 30S subunit contains one RNA molecule of 16S, and the 50S contains two, of 5S and 23S. Antibiotics such as streptomycin and chloramphenicol specifically inhibit protein synthesis by ribosomes.
The relatively higher rate of multiplication in prokaryotic cells than eukaryotic ones is due to the higher number of ribosomes per unit mass. In rapidly growing bacteria, the ribosomes may make up about 40% of the cell’s dry weight, chiefly as polysomes.
Nuclear body. It is of primitive type and also referred to as nucleoid. It is possible to use coloured or fluorescent stains to delineate a central DNA-rich region in bacterial cells. From a lysed protoplast it can be isolated as a structure consisting of loops of supercoiled DNA together with about 10% RNA and 10% protein (mostly RNA polymerase). This nuclear body, an amorphous, lobular mass of fibrillar, intensely chromatinic material occupies about 10-20% of the cell volume. In the electron microscope, this body shows a diffuse area containing fibrous material with no limiting membrane.
Double-stranded DNA forms a single circular chromosome, usually 1-2 mm in length in each nucleoid. The single molecule is about a thousand times longer than the cell itself. Thus, the thread is highly folded to give the bundles of fibres seen in thin sections. The chromosome of E.coli is best studied. This is by far the largest molecule to occur in any biological system.
It is thus hard to conceive the behavior of a molecule that is about one thousand times larger than the largest protein and that exists, moreover, coiled inside the cell several hundred times than itself. Unlike eukaryotes, there are no histones in the nuclear material. However, recently small basic proteins have been detected.
These have been given different names as HU, NS, and DNA-binding protein II. They consist of two very similar polypeptide chains, whose function is said to be similar to eukaryotic histones. The nuclear body exhibits no mitotic or meiotic phenomena. There is no nuclear membrane. Instead, there is an attachment between a specific point on the chromosome and the cell membrane at which replication starts.
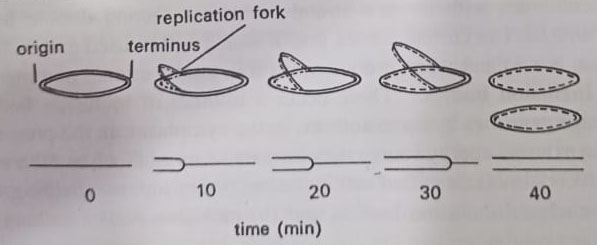
The first stage in nuclear division involves duplication of this attachment site, followed by a progressive bidirectional replication of the DNA by two replication forks, which meet at a point called the terminus. At each replication fork, the nucleotides are added to each naked single strand. In E. coli growing at 370C replication of this length of DNA, almost 1000x the cell length, takes only 40 minutes. This part of the cell is called the C-period. The resulting two daughter chromosomes are then drawn apart by the separation of these attachment points and are induced to coil up to form the two new nuclear bodies.
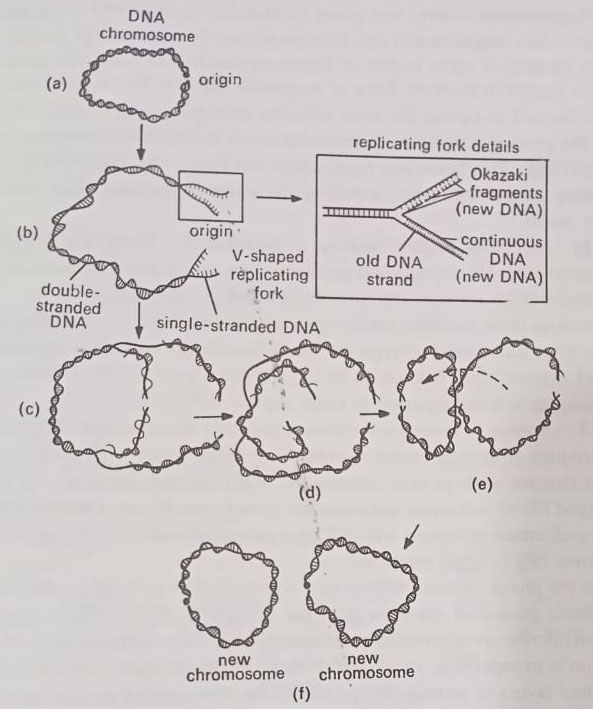
Chromosome replicates by semiconservative method, shown in 1958 in E.coli by Meselson and Stahl. The semi conservative method accounts for DNA replication, but it does not shed light on how a closed-loop chromosome is replicated. This problem perplexed microbiologists until 1962 when John Cairns and his co-workers clarified some of the details of the process.
Cairns, experiments showed that DNA unwinds at a fixed point, whereupon an enzyme nicks the closed loop at a site called the origin. The two strands now separate, or “unzip” to establish a V-shaped replicating fork, as displayed. Synthesis of DNA then occurs along the sides of the fork. In later years microbiologists established that along one side of the fork DNA is manufactured by the continuous assembly of nucleotides, beginning at the origin. (BSc 2nd Year Structure of Microorganisms in Microbiology Notes Study Material)
However, along the other side, the synthesis begins at the point of forking and proceeds in a discontinuous fashion. Here the DNA is synthesized in a series of segments that later join with the help of an enzyme called DNA ligase. The segments came to be known as Okazaki fragments, after Rejii Okazaki who discovered them in 1968. Each of the new DNA strands then combines with a parent strand as Meselson and Stahl postulated. (BSc 2nd Year Structure of Microorganisms in Microbiology Notes Study Material)
The second type of DNA replication is called the rolling circle mechanism. This process takes place in bacteria undergoing the sexual mating process of conjugation. While one strand of DNA remains in a closed loop, an enzyme nicks the other strand. The broken strand then “rolls off” the loop and serves as a template for the synthesis of a DNA strand complementary to itself. When the two strands combine, double helix reforms again. Meanwhile, the intact loop revolves 360 degrees and serves as a template for a strand of DNA complementary to itself.
The loop combines with its new strand to form a second double helix. The bacterium now has two chromosomes, one of which will be used in mating. Sexual reproduction is rare and when it does occur it is unidirectional and incomplete.
Inclusion bodies. There occur a number of inclusion bodies, also called storage granules by some authors, in the cytoplasm. In the presence of an excess of an external energy source, they may make up as much as 50% of the dry weight. Shively (1974) classified cell inclusion bodies into two major groups: (a) membrane-enclosed inclusion bodies, and (b) inclusion bodies lacking a membrane.
(a) Membrane-enclosed inclusion bodies. These are enclosed by a non-unit type membrane, that is 2-4 nm thick and perhaps made up entirely of protein. These inclusion bodies include chlorosomes, carboxysomes, magnetosomes, gas vacuoles, poly-hydroxybutyrate (PHB) granules, sulfur globules, and certain polyglucoside (glycogen) granules.
PHB granules occur in many microbes, including cyanobacteria. Sulphur globules are found in Thiorhodaceae and glycogen in Clostridium spp.
Chlorosomes were demonstrated by Cruden and Stanier (1970) in photosynthetic bacteria of Chlorobiaceae. These differ from chromatophores in being non-membranous, being simply enclosed by nonunit proteinaceous covering. They contain photosynthetic pigments.
Carboxysomes have been demonstrated in chemolithotrophic bacteria and cyanobacteria by Murphy et al (1974). They are said to be involved in the regulation of carbon metabolism.
Magnetosomes-term was given by Balkwill et al (1980) in the bacterium, Aquaspirillum magnetotactic. Blakemore and Frankel (1981) studied magnetotactic bacteria in water bodies of Tasmania and Newzealand. The cells of these bacteria contain iron in the form of magnetite (Fe304). The intracellular magnet allows the cell to pursue the most efficient aerotactic behavior i.e. orientation along the geomagnetic lines of force eliminates the need for twiddling, long runs, and short runs.
This behavior represents a mechanism for migrating down to and remaining in the sediments, avoiding the more oxygenated surface water.
(b) Inclusion bodies lacking a membrane. These are polyglucoside (glycogen) granules, crystals, and paracrystalline arrays and inclusions specific for cyanobacteria i.e. cynophycin granules and phycobilisomes.
Besides those included under types (a) and (b), there are also other inclusion bodies. One such common type is the intracellular refractile structures (R-bodies), demonstrated in bacteria by Lalucat and Mayer (1978). R-bodies are said to be similar to those reported in killer strains of Paramaecium.
Spores. Some prokaryotes, especially Bacillus and Clostridium species produce structures called spores. They have much resistance to environmental stresses such as heat, desiccation, and radiation. They are the means of prolonged life of cells under unfavorable growth conditions. Characteristically, a single endospore is formed within a vegetative cell and on germination, a single vegetative cell is again produced.
In the phase contrast microscope, spores appear as highly refractile bodies sometimes greater in diameter than the cell itself. Thin sections show under an electron microscope a complex multilayered wall. The outermost layer of the variable structure is the exosporium, inside which there is a spore coat composed of several laminated layers of protein. Below this is the thick cortex containing specific peptidoglycan and, below that the protoplast containing the most characteristic chemical composition of the spore – a complex of calcium and dipicolinic acid which perhaps provides heat resistance.
Spore formation occurs after a cell has grown and matured for some hours as a vegetative cell. In Bacillus spores are commonly formed in the center of the cell and are elliptical, whereas, in Clostridium, they are more oval in shape and often formed at the end of the cell or midway between the center and end. Under conditions of a limited supply of C, N, or P, certain Gram-positive rods (aerobic Bacilli and anaerobic Clostridia) and a few sarcinae and actinomycetes, form highly resistant, dehydrated forms, called endospores or spores.
The bacterial spore is perhaps the most resistant living structure known to science due to its multiple coverings. These layers allow it to survive most toxic chemicals and harsh physical conditions. Also, a chemical, dipicolinic acid (DPA) helps to stabilize its proteins. DPA and a large number of calcium ions provide resistance to heat. DPA is liberated as calcium dipicolinate during the germination of spores.
The surrounding mother cell from which spores are eventually released is called the sporangium. Like the seeds of higher plants, they have no metabolic activity. They are particularly adapted for prolonged survival under adverse conditions: they are relatively resistant to killing by heat, as well as by drying, freezing, toxic chemicals, and radiation. Though, resistant to heat, their main ecological role is probably survival in the dry state (or in a non-nutrient medium).
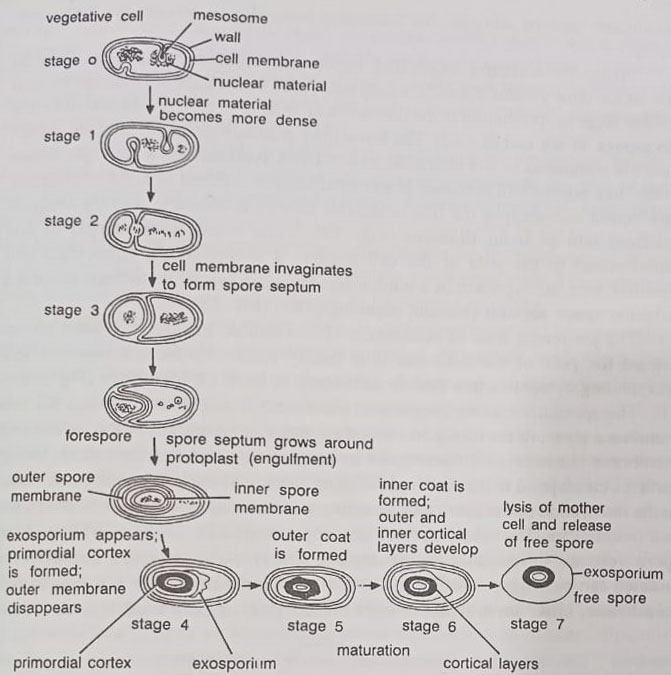
The bacterial spore is not a reproductive structure, as a single vegetative cell forms a single spore and a single vegetative cell will reappear when the spore germinates (cf. a fungus spore which serves both a resistant form as well as reproductive structure). Various stages of the sporulation cycle are shown in Figure. The vegetative cell metabolizes nutrients and multiplies in the vegetative cycle. After some time it enters the sporulation cycle. The overall cycle includes four main stages:
(i) accumulation of cellular DNA to form axial filament and formation of transverse spore septum,
(ii) axial filament is then surrounded by the cell membrane to form a forespore,
(iii) formation of resistant layers of coat material around the spore, completing the formation of the spore, and disintegration of the remaining cell to liberate the free spore, and
(iv) spore germination, when conditions become suitable, the bacterium emerges to resume the vegetative cycle.
Spores are unusually dehydrated, impervious, highly refractile cells. They do not take ordinary stains (Gram’s, methylene blue). In the light microscope, the first visible stage in sporulation is the formation of an area of increased refractility, the forespore at one end of a cell.
The refractility gradually increases and the mature spore is completed in 6-8 hours. Blocks in DNA synthesis show that sporulate starts, like normal cell division, at a critical stage in a round of DNA replication. In electron micrographs, the first detectable change is the conversion of the compact nucleoid into an axial filament. The resulting movement of one chromosome to the pole of the cell triggers a specialized, asymmetrical cell division, with the ingrowth of a double layer of cytoplasmic membrane to form a subpolar spore septum (without peptidoglycan).
The peripheral zone of attachment of this double transverse septum moves toward the pole of the cell, thus finally enclosing the chromosome and surrounding cytoplasm in a double membrane to form the forespore.
The specialized spore integument (envelope) is laid down between the two membrane layers of the forespore, which are initially extensions of the mother cell membrane but become differentiated in composition and function. Both facing surfaces correspond to the wall-synthesizing surface of the parent cell membrane. In the maturation of the spore, a large amount of material is laid down between the two membranes. The resulting thick envelope eventually occupies over half the spore volume, surrounding the protoplast (core). Several layers, successively initiated can be distinguished.
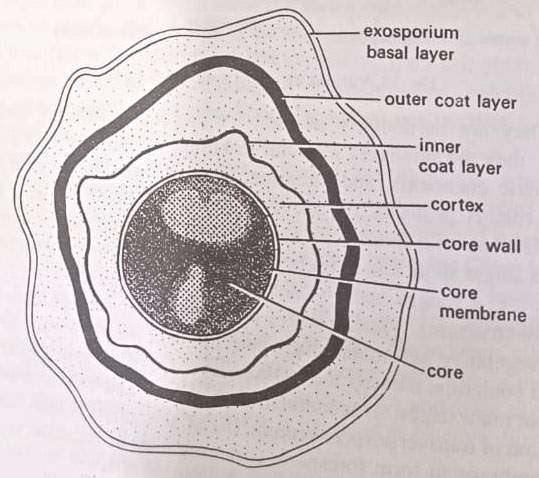
(i) The innermost layer is the germ cell membrane, inner membrane or core membrane, or core wall that surrounds the core.
(ii) Next is the thickest layer, the cortex, which contains a concentrically laminated structure.
(iii) Outside the cortex is the densely stained coat. This may be differentiated into an inner coat layer and an outer coat layer.
(iv) Spores of some species, such as Bacillus thuringiensis are further loosely shrouded in a delicate exosporium or exosporium basal layer.
The cortex contains many layers of spore peptidoglycan, much more loosely cross-linked than that in the vegetative cell. During germination, this part of the peptidoglycan is rapidly autolyzed and released. The coat is made of a keratin like protein, rich in disulfide crosslinks. It constitutes about 80% of the total protein of a spore. The impervious protein coat is responsible for the resistance of the spore to chemicals. The exosporium is a lipid-protein membrane, with 20% carbohydrate. It is not essential for survival, and its function is unknown.
A characteristic feature of the spore is its huge content of Ca2+, normally accompanied by a roughly equivalent amount of dipicolinic acid which can chelate Ca2+. This acid may be as much as 15% of the spore weight. It is located in the core. The core contains much less material than the vegetative cell. It has DNA of one chromosome (3×109 daltons), small amounts of all the stable components of protein-synthesizing machinery (including ribosomes, tRNAs, and accessory factors and enzymes), but no detectable mRNA. Aminoacids (and their biosynthetic enzymes) are absent, supplied early in germination by hydrolysis of a storage protein of low mol. wt, constituting about 20% of the total protein.
The overall process of converting a spore into a vegetative cell is often called germination. It is much faster than sporulation. Three stages can be distinguished; (1) activation-by agents such as heat, low pH, or an SH compound. (2) germination proper or initiation-various metabolites such as alanine, dipicolinate, or inorganic ions such as Mn2+ penetrate the damaged coat and initiate germination. (3) outgrowth- a gradual resumption of vegetative growth, with a progressive increase in protein synthesis and the start of DNA synthesis.
Eukaryotic Microorganisms
They exhibit a tremendous range of structures, functions, behavior, and habitat. Most students would have an idea of the range of these features of eukaryotes from their studies of botany and zoology. It should be clear to them that the structure of eukaryotic cells is generally much more complex than prokaryotic cells. Eukaryotic cells are normally much larger, with a typical diameter ten times greater than that of the prokaryote (i.e. 10 um) and they show much diversity in Size and shape. The yeast cell may be taken as type of eukaryotic cells.
We shall prefer to mention here only some of the components in any detail and an emphasis will be placed on those structures that serve to distinguish the cells fundamentally from those of prokaryotes.
Membranous structures. Within a eukaryotic cell, a variety of membranous selectively permeable structures occur, that gives a complex multi-compartmental whole. Such structures are as follows:
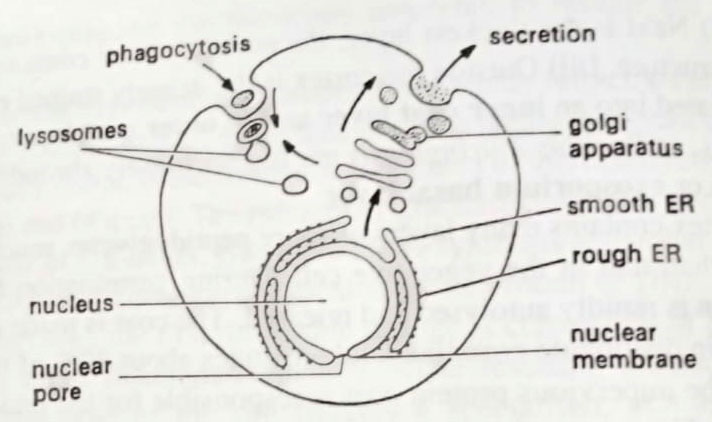
(a) The plasma membrane. In its physical and chemical structure, the plasma membrane is very similar to the cytoplasmic membrane of prokaryotes. However, a difference is the presence of sterols in eukaryotic membranes. In eukaryotes also, this membrane has the same vital function as the prokaryotic cell membrane in being selectively permeable and of transporting specific solutes, but it does not have properties of respiration or photosynthesis. (BSc 2nd Year Structure of Microorganisms in Microbiology Notes Study Material)
An additional feature, however, absents in prokaryotes, can be the ability of wall-less cells to ingest food in particulate form by phagocytosis or in liquid form by pinocytosis, in either case, a membrane-enclosed vacuole is formed within the cytoplasm. Such endocytosis is lacking in bacteria.
(b) Endoplasmic reticulum (ER). The ER typically comprises more than half of the membrane in the cell. It is actually a sac, with the cytoplasm outside and its lumen inside. Its ramifications through the cell give a large surface area. In an electron micrograph two forms of ER is distinguished.
Rough endoplasmic reticulum. It is studded with ribosomes on its outer surface. This is the site of the synthesis of proteins destined for export from the cell, or to become wall components or to be stored in vesicles or vacuoles within the cell, such as the lysosomes which contain digestive enzymes. As they are synthesized these proteins are secreted into the lumen of the ER, often becoming glycosylated to glycoproteins by membrane-bound enzymes.
Smooth endoplasmic reticulum. This lacks ribosomes. It is the site of phospholipid and sterol synthesis in the membrane, and the site of the formation of membrane vesicles which are transferred to the Golgi apparatus for further processing.
(c) Golgi apparatus. This is a series of membranous sacs, originating from the ER. It is a major site of biosynthesis of polysaccharides that are destined for export from the cell, such as pectin-like wall components in algae. Proteins and glycoproteins formed in the ER are covalently processed, for example by further glycosylation, while in the Golgi apparatus. The outer sacs of the Golgi body then release these products in vesicles, some of which move to the plasma membrane, fusing with it, releasing their contents, and adding to its surface area. Other vesicles are stored in the cell, for example, digestive lysosomes.
(d) Nuclear membrane. The nucleus is surrounded by a membrane which is in continuity with the ER and is thus topologically a membranous sac enclosing lumen. The inner-facing membrane has specific proteins on its surface which are said to hold the shape of the nucleus and bind specific regions of chromatin. The nuclear membrane has pores, allowing the transport of molecules in and out of the nucleus. Thus mRNA must be transported out and enzymes such as RNA polymerase must be transported into the nucleus.
(e) Vacuoles, lysosomes, and peroxisomes. As mentioned above, phagotrophic microbes such as many protozoa engulf food particles by invagination of the plasma membrane. Digestion takes place by the fusion of a lysosome with this food vacuole. Vacuoles are formed in many cells. For example yeast, and appear to be involved in the accumulation and storage of metabolic intermediates and ions. The contractile vacuoles occurring in some cells, e.g. freshwater protozoa, function in osmotic regulation and in the excretion of waste products. Peroxisomes are membrane-bound packages of oxidative enzymes such as catalases. They are formed by budding off from smooth ER.
Thus, the above-mentioned membrane systems must be thought of in terms of their spatial and temporal relationships; they are common parts of a dynamic system of membrane flow and modification.
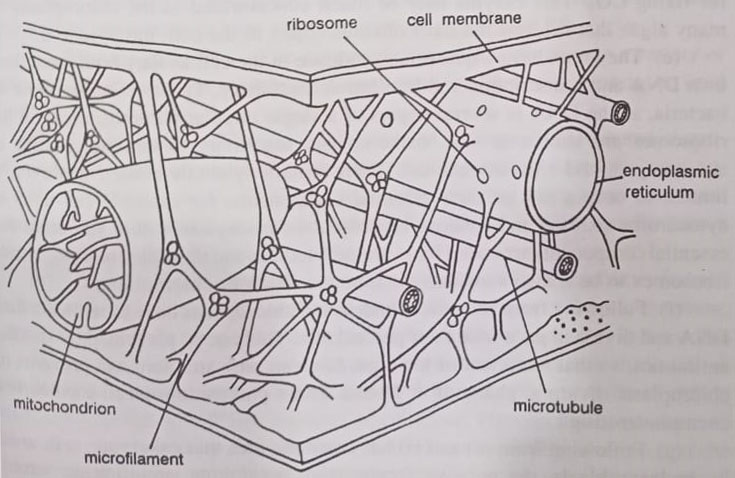
(f) Cytoskeleton. In addition to several membrane-bound organelles, the eukaryotic cell has a cytoskeletal network, consisting of microtubules and microfilaments, that help determine the ability of the cell to move and maintain its shape. This network links the various components of the cytoplasm into a unified structure, the cytoplast, providing the rigidity needed to hold the various structures in their appropriate locations. The network runs throughout the cell, connecting membrane-bound organelles with the cell membrane.
This provides support and movement to organelles, including the cell membrane. It provides the basis for membrane movement involved in the transport of materials into and out of the cell by cytosis. The absence of a cytoskeleton in the prokaryotic cells does not allow cytosis.
Mitochondria and chloroplasts. These two organelles have many features in common, which are as follows:
(a) Both are concerned with energy conversions. Mitochondria are the site of oxidative phosphorylation, converting the energy of oxidative reactions into useful forms, especially ATP. Chloroplasts are the site of photophosphorylation, utilizing light energy to produce ATP and NADPH2, and using these to fix CO2 to carbohydrates.
(b) Both are surrounded by two membranes. The outer one is relatively permeable and resembles the cell membrane, for example by having sterols. The inner one is impermeable and resembles the bacterial cell membrane.
(c) Thus inner bioenergetic membranes have a large surface area. In the mitochondrion, this is the inner membrane which is extensively infolded to form cristae on whose surface are the respiratory chain enzyme complexes. In the chloroplasts within the inner membrane are stacks of membranous sacs (thylakoids) bearing the photosynthetic pigments and associated acceptors.
(d) The inner membrane encloses the matrix. This matrix in each case contains many enzymes; for example in mitochondria those for the TCA cycle, and in the chloroplasts the ribulose biphosphate carboxylase. The enzyme is responsible for fixing CO. This enzyme may be much concentrated in the chloroplasts of many algae that it forms the most obvious object in the cell, the pyrenoid.
(e) They both have quasi-independence in the cell as they both have their own DNA and transcription and translation machinery. These resemble those of bacteria, as the DNA is several copies of a single circular chromosome. And the ribosomes are similar in size, makeup, and sensitivity to antibiotics such as streptomycin and chloramphenicol. Their protein synthetic ability, however, is limited to only a few of their essential components. For example sub-units of cytochrome oxidase and ribulose biphosphate carboxylase. Other sub-units and essential components are coded for by the cell nucleus and translated on cytoplasmic ribosomes to be transported into the mitochondrion or chloroplast.
(f) Following from (e), mitochondria and chloroplasts have to replicate their DNA and divide in pace with their parent cells. This can be prevented by specific antibiotics. So the treatment of the alga, Euglena with streptomycin prevents its chloroplasts from dividing, changing it from a green photoautotroph to a colourless chemoheterotroph.
(g) Following from (e) and (f) has come to the idea that eukaryotic cells arose by endosymbiosis. The putative fermentative prokaryote engulfs an aero bacterium and then a blue-green bacterium. This hypothesis is of course unprovable. But circumstantial evidence comes from the observation that endosymbiosis is a common phenomenon. For example, some strains of Paramecium contain symbiotic bacteria and others contain symbiotic algae which divide in pace with their hosts.
The cytoplasm. The eukaryotic cytoplasm, like that of bacteria, contains many enzymes, metabolites, and other solutes. But is also structured as it contains a cytoskeleton composed of microtubules, actin, myosin, and other structural proteins. Cytoplasmic streaming is common, as seen by the mass movement of organelles within cells.
The nucleus. The nucleus is a definite structural entity surrounded by a membrane and containing many chromosomes. The individual chromosome is made up of linear molecules of DNA compared with the circular DNA of prokaryotes. Associated with the nucleus there is generally an RNA-containing body- The nucleolus which is a specialized structure responsible for ribosomal RNA synthesis.
A function carried out by the single chromosome of the prokaryote nucleus. The eukaryotic chromosomes also differ in having their DNA complexed with basic proteins called histones to form chromatin. As the chromosomes contain much more DNA than the prokaryotic chromosome. Each is replicated by many replication forks, acting in pairs from many sites of origin. This DNA doubling is followed by the complex process of mitosis designed to ensure an orderly partition of a complete set of chromosomes to each daughter cell.
Sexual reproduction is common in eukaryotic microorganisms, and the consequent doubling of the haploid chromosome number then requires meiosis. This allows organisms to have alternate haploid and diploid life cycles.
Storage granules. The eukaryotic microbial cells contain a variety of inclusion bodies such as membrane-bound granules of starch, protein, or lipid droplets.
The cell wall. There is much variation in shape, thickness, and chemical composition of the cell walls of eukaryotic microorganisms. Some protozoa apparently do not have walls. Their plasma membrane may have some additional strengthening to maintain cell shape and rigidity. The cell walls usually have a simpler structure than those of prokaryotes.
(a) Algae. The basic structure is usually maintained by microfibrils formed by intertwining of long polysaccharide molecules, such as cellulose, mannans, xylans, and pectins. In some algae, there is silica or calcium carbonate in the walls.
(b) Fungi. The common structural polymer of the wall is chitin (B 1-4 poly-N-acetyl-glucosamine), often occurring with poly glucose built of B(1-3) ages compared with B(1-4) linkages of cellulose. The mechanical strength of the wall is maintained by cross-linkages between the microfibrils of chitin and the glucan matrix to give a thick and tough structure. Mannans are also common in yeast walls.
(c) Protozoa. There is a great variety of surface components, including structures built from protein, cellulose, calcium carbonate, or silica.
BSc Structure of Microorganisms in Microbiology Notes Study Material
Flagella, cilia, and locomotion. The eukaryotic microbes move usually by the action of flagella and cilia which have a structure quite different from that of the prokaryote flagellum. They are composed of a characteristic “9+2” arrangement of microtubules surrounded by a sheath. Which is an extension of the plasma membrane. They are powered by ATP. Eukaryotic microbes also move by amoeboid movement, as a result of cytoplasmic streaming in cells without a wall. Like prokaryotes, motile eukaryotes show movement towards or away from heat, light, or certain chemical substances. (BSc 2nd Year Structure of Microorganisms in Microbiology Notes Study Material)
Unlike the bacterial flagella that rotate. The flagella and cilia of eukaryotic cells undulate in a wave-like motion to propel the cell. Eukaryotic flagella emanate from the polar region of the cell, whereas cilia, which are somewhat shorter than flagella, surround the cell. Both cilia and flagella are generally involved in cell locomotion. But cilia may also be involved in moving materials, such as food particles.
The movement of flagella or cilia is based on a sliding microtubule mechanism in which the peripheral doublet microtubules slide past each other, resulting in the bending of the flagella or cilia. The peripheral spokes of the microtubular network contain a protein-dynein, which has ATP as activity. And is involved in coupling ATP utilization to the movement of flagella or cilia.
Spores. Eukaryotic microbes form a great variety of spores, some produced in abundance for dispersal, others in small numbers for survival. The latter are generally thick-walled and resistant to harsh environmental conditions.
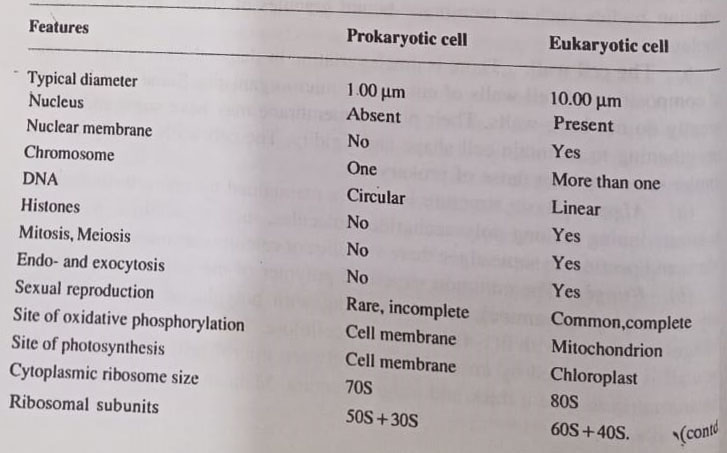

However, the cell chemistry of archaebacteria is very distinct from eubacteria as well as eukaryotes. The numerous and profound differences in organization and function among the three cell types– eukaryotes, eubacteria, and archaebacteria have now become fully recognized. These differences are summarised as follows:


BSc 2nd Year Sample Model Practice Mock Test Question Answer Papers
BSc 2nd Year Structure of Microorganisms in Microbiology Notes Study Material