BSc Survey and Classification of Microorganisms Notes Study Material
BSc Survey and Classification of Microorganisms Notes Study Material: BSc is a three-year program in most universities. Some of the universities also offer BSc Honours. After getting enrolled for BSc, there are certain things you require the most to get better grades/marks in BSc. Out of those, there are BSc Study Material, BSc Sample Model Practice Mock Question Answer Papers along with BSc Previous Year Papers. At gurujistudy.com you can easily get all these study materials and notes for free. Here in this post, we are happy to provide you with BSc 2nd Year Microbiology Survey and Classification of Microorganisms Notes Study Material.

BSc Survey and Classification of Microorganisms Notes Study Material
We shall examine in this Topic the position of microorganisms in the living world and the criteria used in their classification.
Plantae and Animalia
In the early 19th century the microorganisms fitted into the traditional two kingdoms of organisms, the Animalia and the Plantae. This assignment of the microbes to either kingdom was made on the basis of the most easily determinable differences between animals and plants: the ability to active movement and the ability to photosynthesize. Algae (immobile, photosynthetic) were grouped under plants. Fungi were also lumped with the plants. Microscopic motile forms (including protozoa and bacteria) were lumped together as one group, the Infusoria under animals. (BSc Survey and Classification of Microorganisms Notes Study Material)
However, in about 1840 biologists perceived that the Infusoria was a very heterogeneous group in terms of their cellular organization. The simpler, unicellular forms, were, therefore, subdivided into two groups: the protozoa with relatively large and complex cells, and the bacteria with much smaller and simple cells. The old Infusoria was thus split into three groups:
(1) metazoa (multicellular) invertebrate animals,
(2) protozoa (unicellular) animals, and
(3) bacteria, that were transferred to plants.
This Procrustean distribution of microorganisms into plants and animals presented too many inconsistencies. More information became available on microorganisms, that hardly justified their inclusion in either kingdom. For instance, unlike plants fungi and most bacteria are non-photosynthetic. Many bacteria are motile, and some fungi and algae have motile sports. It was thus felt that the division of the living world into these two kingdoms can no longer be maintained.
Protists
A third kingdom, Protists was proposed by E. Haeckel in 1886 to include protozoa, algae, fungi, and bacteria. This kingdom thus included both photosynthetic and non-photosynthetic forms, some plantlike, some animallike, and some having their own specific properties as well as sharing those of both, plants as well as animals. What then distinguished all protists from plants and animals is their relatively simple biological organization. (BSc Survey and Classification of Microorganisms Notes Study Material)
Eukaryotes and Prokaryotes
The development of the electron microscope and some other techniques in 1950 could provide details of the fine structure of cells. Within a few years, there were revealed some hitherto unknown interesting features of cell ultra-structure. This led to a fundamental dichotomy among the various groups of organisms with respect to the internal architecture of the cell: two radically different kinds of cells exist in the contemporary living world:
(1) the more complex eukaryotic cell is the unit of structure in plants, metazoan animals, protozoa, fungi and algae, and
(2) the less complex prokaryotic cell is the unit of structure in two microbial groups: bacteria and cyano-bacteria or blue-green bacteria (formerly known as blue-green algae). (BSc Survey and Classification of Microorganisms Notes Study Material)
The microorganisms thus belong to both these groups. Thus algae, fungi, and protozoa are eukaryotes and all bacteria including cyanobacteria are prokaryotes. Some authors call eukaryotic microbes higher protists and prokaryotic microbes lower protists. If the term protists are still to be retained at all, this has been restricted to the eukaryotic microbes i.e. algae, fungi, and protozoa just to distinguish them from the Plantae and Animalia. (BSc Survey and Classification of Microorganisms Notes Study Material)
However, the chief points of differences between the two will be briefly indicated here. The eukaryotic cells contain a nucleus with a nuclear membrane, multiple chromosomes, and a mitotic apparatus to ensure the equal partition of the products of chromosomal replication to the two daughter cells. The “nucleus” of a prokaryotic cell is a single, circular chromosome, without a nuclear membrane and without histones.
Prokaryotic cells also lack membrane-bound organelles (mitochondria, lysosomes). There is no endocytosis and intracellular digestive vacuoles. They lack (with few exceptions) steroids. They have unique components (muramate and often diaminopimelate) in cell walls. Some prokaryotes are able to fix atmospheric nitrogen and carry obligate anaerobiosis. Some major differences between prokaryotes and eukaryotes. (BSc Survey and Classification of Microorganisms Notes Study Material)
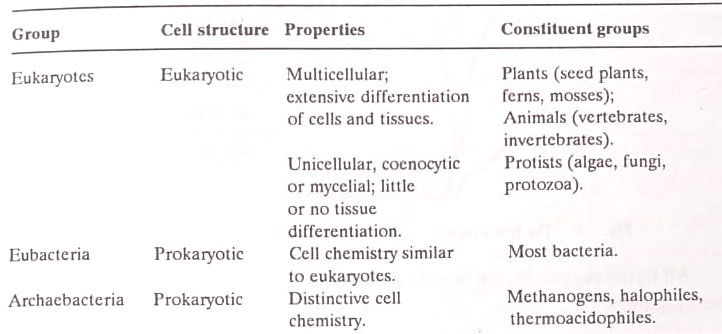
Thus with respect to the internal architecture of the cell, the more complex is the eukaryotic cell, which is the unit of structure in plants, metazoan animals, protozoa, fungi, and algae. The less complex is the prokaryotic cell, the unit structure of two microbial groups: the eubacteria (including the cyanobacteria, formerly known as “blue-green algae”) and the archaebacteria, a heterogeneous group of microbes with prokaryotic structure but with cell chemistry that is strikingly different from that of the eubacteria.
The differences between archaebacteria and eubacteria are so profound that most now believe that this distinction reflects an evolutionary divergence as fundamental as that which divides the eukaryotes from either of the two groups of bacteria.

These newly recognized lines of demarcation run through Haeckel’s proposed kingdom of Protists. Protozoa, fungi, and algae (with the exception of blue-green algae) are eukaryotes that share with plants and animals a common cell structure (eukaryotic) and many details of cell chemistry and function. The eubacteria include most bacterial groups (including the cyanobacteria).
The archaebacteria include only three known groups indistinguishable from the eubacteria on structural grounds but profoundly different chemically. In the future, more groups of archaebacteria could be recognized.
We can thus distinguish on the basis of cell structure and function three major groups of cellular organisms i.e. eukaryotes, eubacteria and archaebacteria as follows :
Monera
In 1969 R.H. Whittaker proposed a five-kingdom system of classification of the living world (Figure 2). He proposed a separate kingdom, Monera for the true prokaryotes: the bacteria and cyanobacteria. Higher protists were distributed among the two kingdoms. Algae and protozoa were grouped under a separate kingdom, Protista, whereas Fungi were given the rank of an independent kingdom. The two other kingdoms were the Plantae and Animalia for traditional plants and animals respectively.
The following Table outlines the changing positions of microorganisms in the living world over the centuries. Viruses are not included as they are entirely a different group.


All living organisms are best thought of today as being members of one of the five kingdoms:
- Monera (prokaryotes, bacteria). This includes Eubacteria (including rickettsiae, chlamydiae, and mycoplasmas). Archaebacteria, and the Oxyphotobacteria (i.e. oxygen-yielding photosynthetic blue-green bacteria, also called cyanobacteria).
- Protista (eukaryotes). It consists of algae, protozoa, slime molds, and water molds. The slime molds and water molds (including other oomycetes) have traditionally been studied by mycologists, and so are usually considered fungi, but phylogenetically they are said to be more related to protists.
- Fungi (eukaryotes). It consists of zygomycetes, ascomycetes, basidiomycetes, and Deuteromycetes.
- Plantae (eukaryotes). The bryophytes and tracheophytes.
- Animalia (eukaryotes). The invertebrates and vertebrates.
Of these, the first three kingdoms (together with the viruses) are studied in microbiology
Main Groups of Microorganisms
There exists a great diversity and complexity among microorganisms. We shall now briefly consider the salient features of the main groups of microorganisms.
[I] Bacteria
They are the true prokaryotes. This group includes the true bacteria (eubacteria) and related organisms i.e. archaebacteria, blue-green bacteria (cyanobacteria or oxyphotobacteria), mycoplasmas, rickettsiae, and chlamydiae.

They are prokaryotes. Reproduction is by simple fission. Nutritional processes are very diverse. Many can thrive in both oxygen-rich and oxygen-free environments. Growth occurs over a wide range of temperatures, usually at a neutral pH. They occur in almost every terrestrial and aquatic environment. (BSc Survey and Classification of Microorganisms Notes Study Material)
Archaebacteria have the following features in common whereby they differ from other bacteria:
(1) Their 16s rRNA molecules are similar to each other but differ greatly from those of other bacteria and from eukaryotes.
(2) Their walls do not contain peptidoglycan, but a range of other unique polysaccharides.
(3) Their cell membrane is of a single layer of glycerol-hydro-carbon-glycerol chains instead of a bi-layer of phospholipids arranged tail to tail.
(4) Their ribosomes are insensitive to chloramphenicol.
(5) They inhabit extreme environments. They include the methanogens (the methane-generating bacteria of anaerobic muds); the extreme red halophiles; the salt-loving bacteria of saturated brine and salted fish; and the thermoacidophiles, found in hot sulphur spring or smoldering coal wastes, It is suggested that they represent a very ancient lineage which diverged from the eubacteria very early in the evolutionary process, and have survived only in these specialized ecological niches.
Rickettsiae, chlamydiae, and mycoplasmas also are prokaryotes. Rickettsiae and chlamydiae are intracellular parasites of eukaryotes. The rickettsiae are tiny rods that are transmitted by arthropods and that multiply only in living cells. Chlamydiae are among the smallest bacteria which were formerly considered viruses. The mycoplasmas are the smallest organisms that can be cultivated outside living tissues. Some are involved in lung disorders. (BSc Survey and Classification of Microorganisms Notes Study Material)
There are some intracellular parasites of prokaryotes. This is a group of small, highly motile bacteria, that adhere to the wall of the host and penetrate through to the periplasm where they replicate, causing eventual lysis of cells. Organisms of this type are called bdellovibrios and perhaps occur in soil.
The blue-green bacteria are typical prokaryotes. They evolve oxygen during photosynthesis and have chlorophyll, a pigment also found in all algae. They are very ancient. They are perhaps major primary producers in the world’s oceans, and many are ecologically important as fixers of atmospheric nitrogen.
Myxobacteria (also called fruiting bacteria) form fruit bodies and specialized multicellular structures like slime molds. Some cells at the tips of the fruit body produce cysts. They are most complex in behaviour and life cycle patterns found in prokaryotes.
[II] Actinomycetes
The majority of them are mycelial. They are Gram-positive prokaryotes, a large group of filamentous bacteria that characteristically show branching patterns just like those of fungi, to give rise to a spreading mycelium. Also, like fungi, they often readily produce spores when grown in culture. They are easily isolated from the soil. Most of these, particularly Streptomyces, produce antibiotics, and so are of great value to the pharmaceutical industry. Their main characteristics are:
(1) Mycelial (majority), Gram-positive prokaryotes.
(2) Some (actinomycetes) develop only in mycelial state reproducing by unicellular specialised spores (e.g. Streptomyces, Micromonospora, Actinoplanes, Streptosporangium, Thermoactinomyces; in others (actinomycetes) mycelial development is transitory and often limited, specialized spores not produced and reproduce primarily by fragmentation into short rod-shaped cells (e.g. Bifidobacterium, Actinomyces, Mycobacterium, Nocardia, Geodermatophilus); and the rest are certain Gram-positive.
Unicellular bacteria which do not form endospores and which are distinguished from the coryneform group by their regular cell shape, and include the lactic acid bacteria (Streptococcus, Leuconostoc, Pediococcus, Lactobacillus) and micrococci (Staphylococcus, Micrococcus, Sarcina).
(3) Cell wall has some peptidoglycan as in nearly all Gram-negative prokaryotes. However, a very large number (almost 60) of other peptidoglycan types have been found in these organisms. The chief variations are the nature of the diamino acid in position 3: the presence, number, and nature of additional amino acids which form interpeptide bridges, and the position of the cross-link between peptide chains.
[III] Protozoa
They are commonly defined as unicellular, eukaryotic animals. To a microbiologist, however, it is not a very helpful definition. It would perhaps be more appropriate to say that they are a group of unicellular non-photosynthetic, eukaryotic microorganisms which normally obtain their food by phagocytosis and which possess no true cell wall. They are involved in many blood and tissue diseases. Movement is an important aspect of the life cycle. A few of their characteristics are as follows:
(1) The type of movement, used to divide protozoa into major groups :
(a) Amoeboid motion as in Amoeba and similar forms.
(b) Flagellar movement as in flagellate protozoa, some of which are the colourless counterparts of particular algae. Some trypanosomes (cause of sleeping sickness) have a simple flagellum, while others as Trichonympha (an organism inhabiting the guts of termites where it is responsible for the wood cellulose eaten by insects) there is a very complex flagellar arrangement.
(c) Ciliary movement in ciliates, as in Paramecium, whose surface is covered with cilia, that are shorter than flagella and have a coordinated motion so that waves of contraction pass over them.
(2) Food, taken by phagocytosis of solid particles such as bacteria.
(3) They have generally no cell wall.
(4) Life cycles are less complicated than algae or fungi, the exceptions being forms like Plasmodium.
(5) Degree of specialization in single cells, as in Paramecium.
Some authors also study slime molds and water molds along with protozoa, thus preferring to treat them as protists rather than fungi. They have been traditionally studied by mycologists. However, evidence is accumulating to show that phylogenetically they are protists. The slime molds form like fungi, macroscopic fruit bodies. But their feeding phase is amoeboid. They live on the surface of decaying vegetation.
There are two main groups: (a) the cellular slime molds, whose vegetative stage is single amoeboid cells that may aggregate to form a pseudo-plasmodium as in Dictyostelium, and (b) acellular slime molds, where a single amoeba produces a multinucleate plasmodium of indefinite size and shape which moves over the surface of the substratum engulfing food particles as it goes.
The water molds are of two main types, the oomycetes and the chytrids, originally included in the fungal kingdom. They are shown as phylogenetically protists, most obviously because they form flagellated motile cells. However, their nutritional processes and vegetative states appear more like fungi than protists. (BSc Survey and Classification of Microorganisms Notes Study Material)
[IV] Algae
They are pigmented eukaryotes, ranging in size from microscopic forms to giant kelps of marine waters. The chief characters are as follows:
(1) Source of energy is oxygen-producing photosynthesis occurring in chloroplasts. Within the chloroplast, there are found a variety of chlorophylls and carotenoids. Chlorophyll is always present.
(2) Exhibit a wide range of morphological types.
(3) Many are motile, usually by flagella.
(4) Cell wall is characteristic of polysaccharide made of components such as pectin, cellulose, or xylan that is sometimes calcified with calcium carbonate. The Wall of some red seaweeds contains agar gel.
(5) Can produce sexually or asexually and can show very complicated life cycles.
[V] Fungi
Some of the chief characteristics are as follows:
(1) Heterotrophic eukaryotic microbes obtain their food in a soluble form by uptake through the plasma membrane in a manner similar to prokaryotes.
(2) Have a thick cell wall usually made of polysaccharides nearly always with chitin microfibrils.
(3) Motile stages absent, never form flagella.
(4) Have a typically branched growth or mycelium made up of individual filaments, the hyphae. Some yeasts are unicellular i.e. non-mycelial.
(5) Mycelia may be coenocytic, or septate.
(6) Asexual reproduction by a variety of spores.
(7) Sexual spores are also produced. In some large fruit, bodies are produced. Life cycles may be simple to complex.
[VI] Viruses
They are acellular, infectious agents. Since they have no associated metabolism one can argue whether they are organisms at all. They have no such activity except replication, that too only in the cytoplasm of live cells. They consist of a fragment of nucleic acid (DNA or RNA, never both) surrounded by a layer of protein. They cause important diseases.
Viroids are minute infectious agents of plants that are composed of single naked RNA molecules. The first report of these pathogenic RNA molecules in the early 1970s aroused much interest since it was previously thought that infectious nucleic acids could survive outside a cell only if they are encapsulated in a protein capsid. Viroid RNAs are very unusual: they are circular, uniquely folded and so small that the largest one so far described (CEV: citrus exocytic viroid) is only 371 nucleotides long-about 1/10th of the size of the smallest RNA virus. Viroids so far are known to cause diseases in plants only.
Prions (proteinaceous infective particles) or slow viruses are a group of recently discovered infectious particles. They are associated with certain degenerative diseases of the central nervous system such as kuru, a disease restricted to a few cannibalistic tribes of New Guinea and transmitted by eating uncooked brain, and other diseases such as scrapie of sheep and goats and Crutzfeldt-Jacob disease of humans and animals. Prions appear to be proteins or glycoproteins with no associated nucleic acid of any kind.
Since replication of the prion-protein presumably occurs by the normal process, this implies that these proteins may be able to bring about the production of their own requisite mRNA. If this were so, prions would have a revolutionary impact on molecular biology since they would constitute the only example where genetic information passes from protein to nucleic acid and not the other way around. However, recent evidence shows that scrapie glycoprotein may be a product of a normal host gene, indicating that these infectious agents may be more convenient than thought previously. (BSc Survey and Classification of Microorganisms Notes Study Material)
Classification of Microorganisms
Before considering the various types of approaches to the taxonomy of microorganisms, we would like to consider the characteristics that are used by microbiologists for this purpose. The following are the types of characters used in classification:
- Morphological characters. These concern cell shape and size, staining reactions, presence or absence of spores or reproductive forms, type of motility, etc.
- Cultural characters. These include the cultural requirements for multiplication (e.g. nutrients, oxygen, temperature, etc.) and the way growth occurs in liquid media, particularly in solid media (e.g. colony form).
- Biochemical characters. These include the more specific biochemical characteristics such as metabolic end-products and the presence or absence of a particular enzyme or pathway.
- Serological characters. These concern the nature of the surface antigens as revealed by suitable specific antibodies.
- Molecular characters. These include the sequences of bases in the DNA, GC ratios, and nucleic acid hybridization.
Approaches to Microbial Classification
With changing history of biology microbial classifications have been attempted with different approaches. These are as follows:
- Artificial approach. The earliest systems of biological classification were artificial. Aristotle, during the fourth century B.C., classified living organisms into two kingdoms-the Animals and Plants. This division was simply based upon (i) the ability of active movement and (ii) the ability to photosynthesize. The microbes were accommodated in both these kingdoms.
- Natural approach. In the middle of the 18th century, Linnaeus (1758) devised a system, that was more useful. This system expressed the biological nature of the objects that it classified. There was scanty knowledge of microbes in this system also.
- Phylogenetic approach. The post-Darwinian biologists thought in terms of evolutionary affinities between organisms. In the 19th century, the concept of a natural system accordingly changed to phylogenetic ones. However, this approach needs sound knowledge of fossil records of organisms. We would like to refer here again to Protista. A German, Ernst H. Haeckel separated the unicellular organisms from plants and animals, placing them under a third kingdom, Protista. The fungi, protozoa, bacteria, and algae were included in this kingdom.

A modern system of this type is devised by R.H. Whittaker (1969). We have already referred to this five-kingdom system of the living world. The microorganisms have been included in the kingdoms Monera, Protista, and Fungi in this system.
- Adansonia approach (Numerical taxonomy). This was first suggested by a French biologist, Michel Adanson, a contemporary of Linnaeus, in the 18th century. Here the taxonomic arrangement is based on the quantification of the similarities and differences among organisms. There are used as many characters as possible, each of which is of equal weight. It is assumed that if each phenotypic character is given equal weighting, it should express numerically the taxonomic distances between organisms, in terms of the number of characters they share, relative to the total number of characters examined. The results are fed into a computer so as to define the similarities and differences between microorganisms and thus indicate possible natural groupings.
- New – the macromolecular approach. The above-mentioned approaches are based mainly on phenotypic traits. The growth of molecular biology has now opened up a number of new approaches to microbial taxonomy. For the identification and classification of bacteria, a range of criteria is now used. Some of the criteria are represented in Figure. The shape and size of the organism, its oxygen, pH, and temperature requirements, and laboratory culture characteristics are considered.
Photosynthetic ability, capsule production, staining reaction, spore-forming ability, and type of movement are other characteristics. Biochemical digestion of certain carbohydrates, proteins, and fats is also considered. There are used certain techniques that give insights into the genetic characteristics of organisms. Two kinds of analysis performed upon isolated nucleic acids furnish information about genotype:
(a) DNA base composition. Although DNA base composition may be determined chemically (hydrolyzing DNA sample and separating the free bases) it can be determined more easily by physical methods, that are now mostly used. The melting temperature or buoyant density of DNA (i.e. the temperature at which it becomes denatured, by breakage of the hydrogen bonds holding the two strands together) is directly related to G+C content. Strand separation is accompanied by a marked increase in absorbance at 260 nm, the absorption maximum of DNA.
When a DNA sample is gradually heated, the absorbance increases as the hydrogen bonds are broken and reach a plateau at a temperature at which the DNA has all become single-stranded. At the midpoint of this rise, the melting temperature (Tm) is a measure of the G+C content. The G+C content may also be determined by subjecting a DNA sample to centrifugation in a CsCl gradient, which affords a precise measure of its density. This method can be used because the density of DNA is also a function of the (G+C): (A+T) ratio.
The mean DNA base composition characteristics of the nuclear DNA in major groups of organisms have been studied. In both plants and animals, the ranges are relatively narrow and quite similar, centering about a value of 35 to 40 mole per cent G+C. Among the protists, the ranges are much wider, the widest range occurring among the prokaryotes, in which the range extends from about 30 to 75 mole per cent G+C. It may be seen (Table 2) that the values vary remarkably among bacteria, from about 30 to 70 mole per cent G+C.
Various streptococci, pneumococci, and lactobacilli have a similar value (38 to 40) which have been traditionally grouped together as lactic acid bacteria, because of their characteristic fermentation. On the other hand, Lactobacillus Bifidus, very much different with 56 percent G+C, is now renamed Bifidobacterium. Within other large groups also, a considerable range may be seen which have been grouped into a single genus, mainly on morphological grounds. For example, range in different species of Proteus, Bacillus, and Corynebacterium.

If one examines the mean G+C content of many different strains of a single microbial species, the values are closely similar or identical, as shown for several species of Pseudomonas. Each bacterial species accordingly has DNA with a characteristic mean G+ content, which can be considered one of its important specific characters.
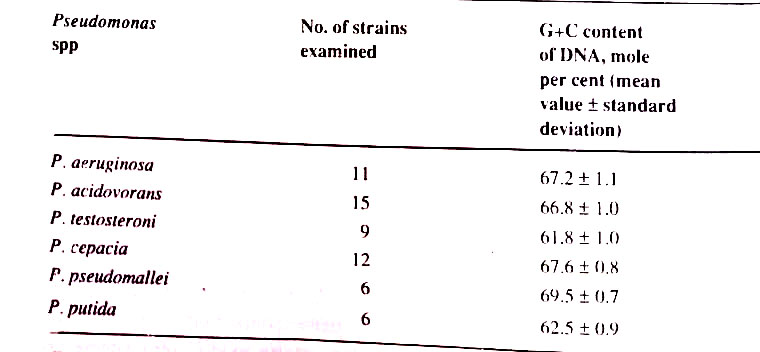
It has been observed in this approach that a substantial divergence between two organisms with respect to mean DNA base composition reflects a large number of individual differences between the specific base sequences of their respective DNAs. It is very important, as the major genetic divergence reflects a wide evolutionary separation. However, this may not be true, since two organisms with similar mean DNA base composition may differ greatly in the genetic constitution. This is true for plants and animals. However, this technique proved useful for microbes. (BSc Survey and Classification of Microorganisms Notes Study Material)
(b) Nucleic acid hybridization. (DNA-DNA homology or DNA base sequence and RNA sequences). The similarity of the base composition represents only a limited basis for close genetic relatedness because even distant organisms can by chance have a similar composition. Moreover, as groups diverge in evolution the DNA sequence changes long before the base composition.
Homology of sequence (DNA – DNA homology) can be measured quantitatively in terms of the ability of DNA strands from two different sources to form molecular hybrids in vitro. Surveys among higher organisms have revealed close parallelism between the results of such hybridization and the phylogenetic relations concluded on other grounds. When DNA preparations from two related strains of bacteria were mixed and treated at 10 to 30°C below the Tm value, hybrid DNA molecules are formed.
However, DNA preparations from two unrelated bacteria, could not hybridize. This discovery of reassociation of single-stranded DNA molecules (single strands formed due to treatment given at a temperature below Tm value) from different biological sources to form hybrid duplexes laid the foundations of DNA-DNA homology in bacterial genetic relatedness.
Among bacteria, this is useful only within closely related groups because it quickly disappears in the wider range of variation. Since duplexes can also be formed between single-stranded DNA and complementary RNA strands, analogous DNA-RNA reassociations can be performed. Ribosomal RNA hybridization to DNA is useful for estimating more distant kinship among bacteria.
RNA sequences have proved more useful than DNA sequences because they involve relatively small segments of the chromosome: those that code the base sequence either of the tRNAs or of the rRNAs. An alternative is thus to sequence the smaller molecules (RNA or protein) derived from DNA. This technique is thus used for the comparative sequencing of specific RNA molecules. The molecules are highly conserved in an evolutionary sense and provide a very useful measure of the phylogenetic relationships of the wider groupings of microorganisms. The chosen molecules are ribosomal RNA components, the 165 rRNA and the 55 rRNA.
However, nucleic acid hybridizations have not yet provided a phylogenetic bacterial taxonomy to replace the classical Bergey’s Determinative key.
Different groups of microorganisms have been classified in different ways. The classificatory accounts may be found in different textbooks on individual groups of the microbial world. The bacteria present a special case.
Main Outline of Bacterial Classification
The recently developed molecular approaches have provided a useful supplement to the classic approaches based solely on phenotypic characters. It is now possible to recognize among the bacteria a considerable number of subgroups that appear somewhat natural.
Many of them could be distinguished by both phenotypic and genotypic criteria. It is still not possible to develop a complete hierarchical classification for bacteria. The earliest recognized system of bacterial taxonomy was devised by David H. Bergey, who first published his system in 1923 in his book, Bergey’s Manual of Determinative Bacteriology.
Since then there have been considerable changes in the composition and arrangement of higher taxonomic groupings. His classification given in the eighth edition (1974) has been accepted as the official document of bacteriologists. In this system, bacteria constitute as one of the Divisions of the kingdom, Prokaryotae. This is given in Table.


His book was updated later and appeared as Bergey’s Manual of Systematic Bacteriology. The first volume of the first edition of this book was published in 1984 and the subsequent three volumes appeared in the following years. The editors placed bacteria in the kingdom Prokaryotae, divided further into four divisions. All bacteria are grouped into 30 sections. Volume I has 11, volume II has 6, volume III has 9, and volume IV has 4 sections. (BSc Survey and Classification of Microorganisms Notes Study Material)
Volume I deals only with Gram-negative bacteria of general, medical, and industrial importance; volume II with Gram-positive bacteria other than the filamentous actinomycetes; volume III with archaebacteria, the cyanobacteria, and the remaining Gram-negative bacteria including the chemolithotrophs; and volume IV covers the Gram-positive, filamentous bacteria. The newest edition of Bergey’s Manual does not use a hierarchical ordering of bacteria; instead, they simply organize the bacteria into groups that have no official taxonomic status.
BSc Survey and Classification of Microorganisms Notes Study Material
BSc 2nd Year Sample Model Practice Mock Test Question Answer Papers